Электрическая активность мозга
В основе электрической активности мозга лежит потенциал действия и синаптические взаимодействия между нейронами. Потенциал действия – это не что иное, как временный сдвиг (от отрицательного к положительному) мембранного потенциала нейрона (-70 мВ), вызванный потоками ионов изнутри и внутрь клетки. Он обусловлен наличием в клеточной мембране особых типов потенциалзависимых ионных каналов. Клетка срабатывает, производя потенциалы действия, который распространяются вниз по ветвям нейронов в виде цепных реакций, вызывая волну деполяризации и реполяризации. Потенциалы действия движутся только в одном направлении.
Таким образом, потенциал действия движется вдоль ветви пока не достигает конца аксона, на котором расположен синапс. Синаптическое взаимодействие может влиять на постсинаптический нейрон как тормозящим, так и возбуждающим образом. Связывание нейротрансмиттеров с рецепторами в постсинаптическом нейроне, либо перетекание заряда через щелевые контакты может вызвать либо кратковременные изменения, такие как изменения мембранного потенциала, называемые постсинаптическими потенциалами, либо более долгосрочные изменения путем активации сигнальных каскадов.
На уровне целого мозга электрическая активность проявляется как ритмические колебания электрического потенциала, наблюдаемые в нейронных ансамблях. Эти колебания, которые существуют в диапазоне частот от одного до нескольких сотен герц, доминируют в электрофизиологических измерениях крупномасштабной активности мозга и образуют мозговые ритмы и волны. Осцилляции можно охарактеризовать их частотой, амплитудой и фазой. Эти свойства сигнала могут быть извлечены при помощи регистрации электрической активности нейронов мозга с использованием частотно-временного анализа. Нейронные осцилляции и синхронизация связаны со многими когнитивными функциями, такими как передача информации, восприятие, контроль движений и память. 1 2 3
Электроэнцефалография
Электрическое поле, создаваемое одиночным нейроном или совокупностью близкорасположенных нейронов, настолько мало, что не может быть зарегистрировано с поверхности головы. Электроэнцефалография или ЭЭГ отражает суммарную активность большого пула нейронов, величина которого достаточна для формирования разницы потенциала на поверхности скальпа.
Следует подчеркнуть, что электрическая активность должна быть настолько большой мощности, насколько это потребуется для прохождения сигнала через кости черепа и кожные покровы, обладающие заметным сопротивлением электрическому току. Опять-таки, исходя из приведенных необходимых и достаточных условий: активность, регистрируемая со скальпа, обусловлена относительно мощной суммарной активностью нейронального пула, — логично вытекает положение о том, что суммарная ЭЭГ в первую очередь обусловлена активностью нейронов коры как самых близко расположенных к поверхности кожи головы — месту регистрации.
Мозговые ритмы и волны
➥ Более подробно: Ритмы ЭЭГ
В центральной нервной системе (ЦНС) нейроны связываются посредством электрохимических сигналов, которые приводят к потоку ионных токов через синаптические контакты. На сетевом уровне синхронизация электрической активности нейрона вызывает ритмические колебания напряжения, распространяющиеся по областям мозга, известные как нейронные колебания или мозговые волны. Они выполняют различные физиологические функции и коррелируют с различными поведенческими состояниями. Мозг обладает множеством разных типов осцилляторов с различными периодами колебаний. Осцилляторы одновременно выдают частоты от 0,02 до 600 Гц. Классификация частотных границ позволила создать классификацию, по которой можно описывать ритмы мозга. 4 На мезо- и макроуровнях эти колебания подразделяются на альфа-активность (8-12 Гц), 5 которая обнаруживается в затылочной доле во время спокойного бодрствования и усиливается при открывании глаз, 6 дельта (1-4 Гц), тета (4-8 Гц), мю (8-13 Гц), бета (13-30 Гц), низкая гамма (30-70 Гц) и высокая гамма (70-150 Гц) частотные диапазоны, которые могут быть измерены с помощью электроэнцефалографии.
Ритмический характер ЭЭГ определяется, по меньшей мере, тремя ритмообразующими компонентами.
- Первый – управляющее влияние ретикулярной формации среднего мозга на таламус; 7
- Второй компонент – таламус и лимбическая система;
- Третий – пирамидные нейроны коры.
Предполагают, что активность коры модулируется пейсмекерами таламуса, которые в свою очередь имеют периферические входы и получают сигналы от ретикулярной формации ствола мозга.
Учеными было установлено, что целесообразной моделью для описания распределения альфа-ритма по скальпу стоит считать двухуровневую модель генерации. Первый представлен двумя базовыми генераторами в ретикулярных ядрах таламуса, второй – двумя модально-специфическими корковыми генераторами, локализованными в определенных специфичным модальности корковых областях. 8
В зависимости от эмоций, чувств и состояния человека в целом всегда преобладает только одна определенная волна. В целом они привязаны к какому-либо виду мышления или ритму деятельности.
Виды мозговых волн
Альфа-волны ритмичные колебания, частотой от 8 до 14 Гц, средняя амплитуда 30-70 мкВ. Регистрируются почти у всех здоровых людей в состоянии спокойной мозговой деятельности при бодрствовании. При этом ритме организм способен производить успокаивающие нейрохимические вещества. Данный ритм выражен в теменных и лобных областях.
На данный момент ученые рассматривают несколько теории происхождения и локализации генераторов альфа-активности:
- факультативная теория пейсмекеров. Изложена П. Андерсоном и его коллегами. Она базируется на локализации центрального механизма генерации альфа ритма в таламических ядрах, влияющих на нейронную активность в соответствующих корковых областях; 910
- модель корковых и таламических генераторов предложенная Ф. Лопесом да Сильва с сотр., 11 которая основана на существовании относительно независимых генераторов, находящихся как в ядрах таламуса, так и в коре;
- теория Басара Е. 12 утверждающая наличие множества мультифункциональных селективно распределенных по мозгу генераторов альфа- ритма, образующих диффузно-распределенную альфа-систему;
- В свою очередь, Williamson S.I. полагает, что в мозговой коре есть самостоятельные автогенераторы, т.е. дискретные источники осцилляций, называя такие источники альфа диапазона «альфонами» . Кроме того, есть предположение, что подобным же образом в коре головного мозга существуют и генераторы бета-, тета-. дельта- ритмов. 13
В последних исследованиях были выделены новые пограничные подгруппы альфа-волн – Мю волны (Тальфа) в диапазоне от 7 до 13 гц. Активизируются при визуализации движений или при физ. активности. Исходят из передних частей полушарий.
Бета-волны – высокочастотные (13-100 Гц), но низкоамплитудные (14-40 мкВ). Их регистрируют в лобных областях в нормальном состоянии бодрствования, мы внимательны и сконцентрированы. При взволнованном или тревожном состоянии генерируются быстрые бета-волны, а при ощущении угрозы и опасности – сверхбыстрые.
Гамма-волны (разновидность бета-ритма) – самые быстрые волны. Находятся в диапазоне 30-170 Гц (в некоторых источниках до 500 Гц). Амплитуда крайне низка – ниже 10 мкВ. Встречаются при решении сложных проблем или задач, описывается состояние «эврики». Так же возможны появления при волнении и страхе.
Тета-волны – медленные волны частотой 4-8 Гц и амплитудой 20-60 мкВ. Эти волны наиболее интенсивны в затылочной и височных областях. В норме регистрируются у детей или при эмоциональном стрессе у взрослых (в подавленном состоянии, фрустрации).
Дельта-волны – все низкочастотные волны на ЭЭГ (ниже 3,9 Гц). Низкоамплитудные (20-30 мкВ). Генерируются при глубоком сне или при коме. У детей могут проявляться при серьёзных органических поражениях мозга. Они считаются «истинно корковыми» волнами из-за их автономности при активности нижележащих областей мозга.
Ритмы головного мозга
Альфа-ритм
➥ Более подробно: Альфа-ритм
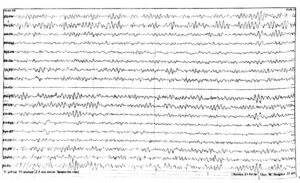
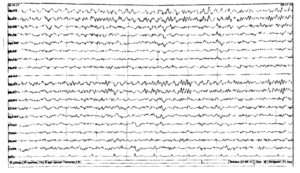
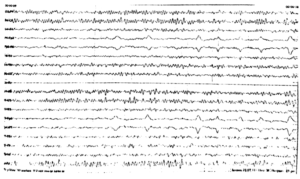
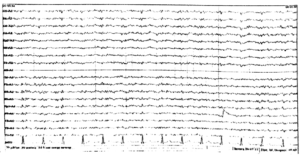
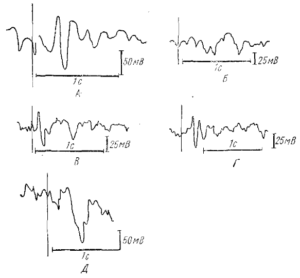
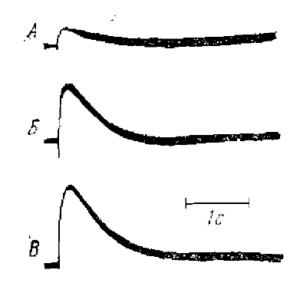
Регистрируется преимущественно в затылочных отведениях в виде синусоидального, модулированного в веретена по амплитуде ритма частотой 8-13 Гц, амплитудой до 100 мкВ. По направлению к передним отведениям его амплитуда и индекс уменьшаются. Наибольшая амплитуда альфа-ритма отмечается в состоянии спокойного, расслабленного бодрствования, в темном помещении, при закрытых глазах. При открывании глаз он редуцируется, а при закрывании глаз восстанавливается (рис. 1-3).
Сенсомоторный мю-ритм
➥ Более подробно: Мю-ритмы
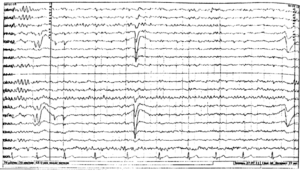
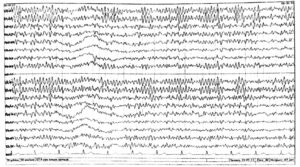
Встречается преимущественно в молодом возрасте. Выявляется на ЭЭГ в центральных отведениях в виде аркообразного ритма, частотой равной или немного превышающей альфа – ритм (рис. 4).
На открывание и закрывание глаз не реагирует. Активируется умственной нагрузкой и психическим напряжением. Редуцируется при активации двигательной активности в конечностях (проба со сжатием кулаков).
Бета-ритм
➥ Более подробно: Бета-ритмы
Регистрируется преимущественно в передних отведениях, частотой 14-40 Гц, амплитудой до 20 мкВ , шумоподобный, не реагирует на открывание и закрывание глаз (рис. 5).
Бета – ритм связан с соматическими, сенсорными и двигательными корковыми механизмами и дает реакцию на двигательную активность или тактильную стимуляцию. При выполнении или умственном представлении движения бета-ритм исчезает в зоне соответствующей корковой проекции.
Тета – колебания
➥ Более подробно: Тета-ритмы
Регистрируются преимущественно в передних отведениях, в виде единичных волн, по амплитуде не превышающих основной корковый альфа-ритм (рис. 6). В отдельно-взятой эпохе ЭЭГ количество тета – волн должно составлять у молодых до 20%, постепенно увеличиваясь с возрастом до 40%.
Нормальная ЭЭГ взрослого бодрствующего человека
У большинства здоровых взрослых в состоянии спокойного бодрствования на ЭЭГ доминирует альфа-ритм, с амплитудным преобладанием в теменнозатылочных отведениях. По направлению к передним (лобно-центральным отделам) альфа-ритм убывает по индексу и амплитуде и сочетается с бета активностью (рис. 6). У 10-15 % здоровых обследуемых, регулярный альфа-ритм не превышает 20 мкВ и по всему мозгу регистрируются высокочастотные, низкоамплитудные колебания, которые свидетельствуют о преобладании в мозге десинхронизирующих влияний. Это тип ЭЭГ связан с аутосомнодоминантным геном. Л. Р. Зенков указывает и на половые различия ЭЭГ: женщинам свойственны более высокие частоты альфа – ритма и большее количество бета-активности. Автор делает вывод, что эти данные определенным образом коррелируют с половыми различиями психологических характеристик, свидетельствуют о более высоком уровне активации у женщин, имеют генетическую обусловленность и могут быть связаны с гормональными особенностями. У некоторых здоровых обследуемых отсутствует нормальный альфа-ритм, а вместо него регистрируется активность частотой 14-18 Гц, амплитудой около 50 мкВ, с преобладанием в затылочных отведениях и снижением амплитуды по направлению к передним отделам (рис.2). При умственной нагрузке, эмоциональном напряжении этот ритм исчезает (реакция активации).
Альфа-ритм выражен в затылочных отведениях, модулирован по амплитуде, частотой 9 Гц. Межполушарная асимметрия отсутствует. Зональные различия сохранены. Бета-ритм преобладает в передних отведениях, невысокой амплитуды легко деформирован тета- волнами. Медленная активность представлена единичными тета- волнами, по амплитуде не превышающими фон, регионарно преобладающих в передних отведениях, без склонности к группированию и фокального акцента. Дельта волны – отсутствуют.
Читайте также
- Пачка спайков / вспышка (Bursting)
- Нейронная активность
- Модель одиночного диполя
- Активность одиночного нейрона
- Активность нейронной популяции
- Локальные потенциалы
- Потенциал действия нейрона
- Вызванные потенциалы
Footnotes
- Fries P (2005). “A mechanism for cognitive dynamics: neuronal communication through neuronal coherence”. Trends in Cognitive Sciences. 9 (10): 474–480. doi:10.1016/j.tics.2005.08.011.
- Fell J, Axmacher N (2011). “The role of phase synchronization in memory processes”. Nature Reviews Neuroscience. 12 (2): 105–118. doi:10.1038/nrn2979.
- Schnitzler A, Gross J (2005). “Normal and pathological oscillatory communication in the brain”. Nature Reviews Neuroscience. 6 (4): 285–296. doi:10.1038/nrn1650.
- Buzsáki, G (2006). The Rhythms of the Brain. Oxford Press.
- Foster, JJ; et al. (July 2017). “Alpha-Band Oscillations Enable Spatially and Temporally Resolved Tracking of Covert Spatial Attention”. Psychological Science. 28 (7): 929–941. doi:10.1177/0956797617699167.
- Berger H, Gray CM (1929). “Uber das Elektroenkephalogramm des Menschen”. Arch Psychiat Nervenkr. 87: 527–570. doi:10.1007/BF01797193.
- Фокина Ю. О., Павленко В. Б., Куличенко А. М.. “ВЕРОЯТНЫЕ МЕХАНИЗМЫ ГЕНЕРАЦИИ ЭЛЕКТРОЭНЦЕФАЛОГРАММЫ”// Ученые записки Крымского федерального университета имени В. И. Вернадского. Биология. Химия, vol. 20, № 4 (59), 2007, pp. 96-108
- Исаичев C.A.. Осипова Д.С.. Коптелов Ю.М. Дипольные модели генераторов альфа-ритма // ЖВНД. – 2003. – Т. 53. № 5. – С. 577-586
- Andersen P.. Andersson S.A. Thalamic origin of cortical rhythmic activity / Handbook of Electroencephalography and Clinical Neurophysiology / Ed. Amsterdam: Elsevier. 1974. – Vol. 2 , – P. 90
- Andersson S. A.. Holmgren E. Theoretical consideration on the synchronization of thalamo-cortical activity Subcortical Mechanisms and Sensorimotor Activities / Ed. Frigyesi T.L. Bern: Huber. 1975. – P. 229-250.
- Lopes da Silva F.H.. van Lierop T.H.M.T.. Schrijer C.F.M.. Storm van Leeuwen W. Essential differences between alpha rhythm and barbiturate spindles: spectra and thalamo-cortical coherence Electroenceph. clin. – 1973b. – V. 35. – P. 626-639
- Basar E. Brain function and oscillations. – Berlin. New York: Springer. 1998. – 467 p
- Лобасюк Б.А. Роль ретикулярной формации ствола мозга в механизмах электрогенеза Нейрофизиология. – 2005. – Т. 37. №1. – С.39-45
Общая классификация различных видов электрической активности головного мозга и их краткая характеристика
Общая классификация электрической активности головного мозга является сложной задачей, в то же время совершенно очевидно, что необходимость такой классификации давно назрела, так как в общей электрофизиологии мозга и клинической электроэнцефалографии списан широкий спектр различных ЭЭГ феноменов и определенное их упорядочивание и классификация необходимы. Здесь возникает ряд трудностей и прежде всего правомочность классификации, казалось бы, сходных ЭЭГ феноменов, описанных у животных и человека, т. к. мы еще не всегда имеем право относить их к одному классу явлений. Другая трудность заключается в том, что на ряд биоэлектрических процессов мозга в литературе часто существуют диаметрально противоположные взгляды, совместить которые не всегда возможно. И, наконец, наибольшие сомнения возникают там, где совершенно не ясен генез тех или иных электрических явлений, а известны только общие внешние характеристики. Поэтому и разбираемая здесь классификация, не будет лишена недостатков.
Предложенная классификация является расширенным и измененным вариантом уже имеющихся классификаций (Соколов, 1962; Кожевников, Мещерский, 1963). Мы остановимся на всех видах суммарной электрической активности мозга.
Всю биоэлектрическую активность головного мозга можно разделить на две крайние группы: импульсную активность и суммарную медленную активность (см. схему общей классификации различных видов электрической активности головного мозга).
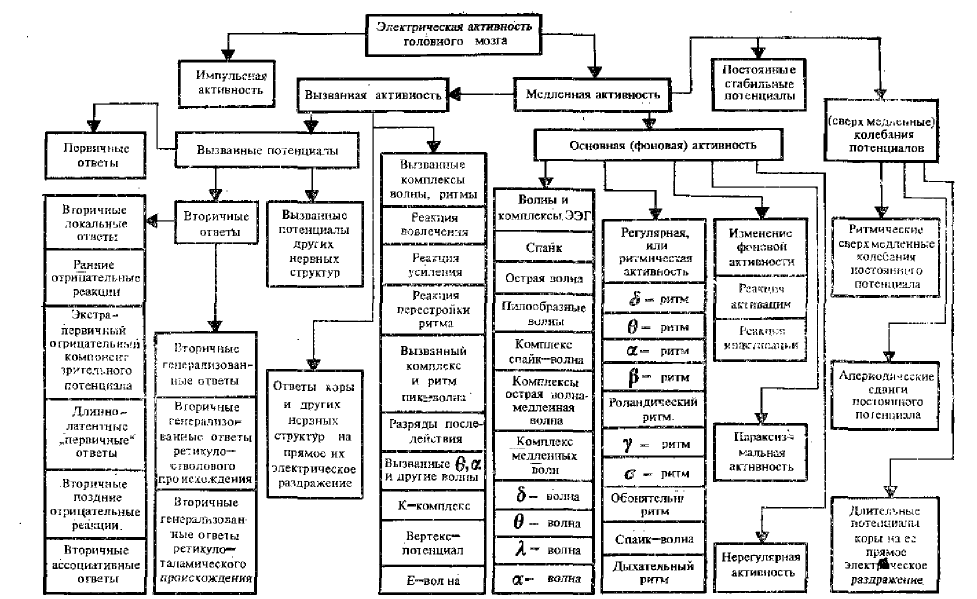
Импульсная активность представляет собой в основном форму деятельности аксонов, тел нервных клеток и частично дендритов.
Медленная активность определяется, как мы уже говорили ранее, в основном алгебраической суммой ВПСП и ТПСП сомы и дендритов нервных клеток, а также, видимо, сдвигами метаболических процессов мозга и активностью глиальных элементов.
Медленная суммарная активность головного мозга делится на три основные, группы:
- Вызванную электрическую активность мозга;
- Основную («спонтанную», фоновую, автономную) активность;
- Медленные и сверхмедленные колебания потенциала.
К вызванной электрической активности головного мозга относятся простые и сложные (комплексные) колебания биопотенциалов, возникающие в коре и различных других образованиях головного мозга при прямом электрическом, одиночном или ритмическом их раздражении и в ответ на поступление в них импульсов по различным афферентным путям при адекватной или электрической стимуляции. Они объедины под общей рубрикой — вызванные потенциалы.
К вызванным потенциалам относятся:
- Первичные ответы коры больших полушарии;
- Вторичные ответы коры больших полушарий;
- Вызванные потенциалы других нервных центров (например, гиппокампа, мозжечка, таламических структур и т. д.);
- Ответы коры и других нервных структур на прямое их электрическое раздражение.
Кроме того, к вызванной активности отнесены спровоцированные различными способами комплексы, волны и ритмические электрические процессы, которые в норме характерны для спонтанной суммарной активности того или иного отдела мозга.
К вызванным комплексным реакциям, волнам и ритмам относятся:
- Реакция вовлечения;
- Реакция усиления;
- Реакция перестройки ритма;
- Вызванный комплекс и ритм пик-волна;
- Разряды последействия;
- Вызванные θ-, α-, обонятельные волны и т. д;
- К-комплекс, вертекс-потенциал, Е-волна (волна «внимания»).
Дадим краткую характеристику всем видам активности.
Первичные ответы коры больших полушарий. К ним относятся ответы, возникающие только в корковых проекционных зонах анализаторов животных и человека при сенсорных адекватных раздражениях или при электрическом раздражении рецептора, проводящих специфических путей и релейных ядер.
Вторичные ответы коры больших полушарий возникают при адекватных и неадекватных раздражениях рецептора, а также определенных групп таламических ядер и ретикулярной формации ствола мозга.
Электрогенез вторичных ответов отличается от такового первичных ответов. Вторичные ответы можно разделить на:
- Вторичные локальные ответы, куда относятся: а) ранние отрицательные реакции, б) экстрапервичный отрицательный компонент зрительного потенциала, в) длиннолатентные «первичные» ответы, г) вторичные поздние отрицательные реакции, д) вторичные ассоциативные ответы, е) вторичные коллазальные ответы;
- Генерализованные ответы, куда относятся: а) вторичные генерализованные ответы ретикуло-стволового происхождения и б) вторичные генерализованные реакции ретикуло-таламического происхождения.
Вызванные потенциалы других нервных центров
Электрогенез этих видов реакций (так же как и в коре больших полушарий) целиком определяется характером афферентных связей и строением центров. Они возникают также на адекватное или электрическое раздражение рецепторов, проводящих путей или центров, с которыми исследуемый центр имеет связи. Так, например, вызванные потенциалы мозжечка возникают на световую, звуковую, сомато-сенсорную стимуляцию, на прямое электрическое раздражение тех же рецепторов, ряда таламических ядер и других подкорковых ядер, а также на прямое электрическое раздражение отдельных зон коры больших полушарий.
Реакция вовлечения (recruiting response) состоит из серии медленных отрицательных волн, возникающих с большим латентным периодом в коре больших полушарий, при ритмическом, низкочастотном (5—10 в 1 с) электрическом раздражении ядер медиального таламуса.
Впервые описана и исследована Демпси и Морисоном (Dempsey, Morison, 1942).
Реакция усиления (augmenting response) возникает в коре больших полушарий при низкочастотном раздражении специфических ядер таламуса и волокон радиации. Описана в 1943 г. теми же исследователями.
Реакция перестройки ритма впервые была описана. Эдрианом и Мэттьюзом (Adrian, Mattews, 1934) в электроэнцефалограмме человека. Они называли ее «реакцией мельканий» (flicker response). В дальнейшем эту реакцию разные авторы стали называть «реакцией навязывания ритма, следования ритма и усвоения ритма». Мы будем пользоваться термином «реакция перестройки ритма».
Эта реакция заключается в изменении, перестройке спонтанной активности коры мозга под влиянием, как правило, световых ритмических раздражений сетчатки глаза. В этом случае электрическая активность коры приобретает ритмический характер с частотой, равной или в целое число раз большей или меньшей по сравнению с частотой применяемого ритмического светового раздражения.
Реакция перестройки ритма наблюдается и при ритмических афферентных раздражениях других модальностей. Например, описана реакция перестройки в ЭЭГ человека при действии ритмического звука (Davis et al., 1939; Xeher, 1961).
В ЭЭГ животных впервые реакция перестройки ритма на ритмический световой раздражитель описана и исследована Бартли (Bartly, 1936) и М. Н. Ливановым (1940).
Комплекс спайк-волна и ритм спайк-волна
➥ Основаня статья: Спайк-волна ЭЭГ
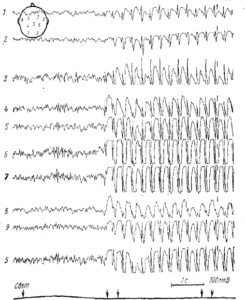
Комплекс спайк-волна является патологическим комплексом в ЭЭГ животных и человека и характерен для специфического случая генерализованной эпилепсии, носящей название «petit mal», или «малого миоклонического припадка» (рис. 1). Характеризуется комбинацией спайка и следующей за ним медленной волны длительностью от 200 до 500 мс. В литературе встречается и другой термин для этого комплекса — спайк-купол (spike-deme). Как правило, во время клинического припадка petit mal эпилепсии в ЭЭГ возникает ритм спайк-волна, который состоит из ритмически повторяющихся с частотой 2,5—3,5 кол/с патологических комплексов спайк-волна. Поскольку наиболее выраженным компонентом комплекса при petit mal —эпилепсии является волна, а не пик, то некоторые исследователи (Кругликов и др., 1970) предлагают назвать комплекс не спайк-волна, а волна-пик.
Комплекс спайк-волна и спайк-волна ритм могут быть вызваны искусственно у животных рядом способов, путем стимуляции неспецифических таламических ядер химически (введением алюминиевой пасты) или с помощью электрического раздражения тех же ядер с частотой 3 в 1 с.
Разряды последействия
Возникают в ЭЭГ при различных одиночных адекватных раздражениях того или иного анализатора путем прямой электрической стимуляции ряда ядер таламуса или коры. Разряды последействия описаны и изучены у животных (Bartly. Bishop, 1933; Adrian, 1941; Bremer, 1949; Chang, 1951; Брейзье,. 1952; Гусельников. Супин, 1962, 1968, и др.) и у человека (Cigancc, 1958; Barlou, 1960; Cohn, 1964, и др.)
Все разряды последействия имеют, как правило, вид вспышки ритмических электрических колебаний в коре (или таламическом ядре) и возникают после начального ответа на применяемое раздражение, например, после первичного ответа в коре в случае афферентного раздражения или дендритного ответа в случае прямого Электрического раздражения коры и т. д. (рис. 2). Обычно вспышка волн ритмического разряда последействия (РРП) носит затухающий характер; вначале амплитуда волны максимальна, к концу постепенно убывает. Частота волн PPП различна у различных животных и человека. У человека она близка к частоте α-ритма (10— 12 Гц), у кошек около 10 Гц (в барбитуровом наркозе). По Эдриану (Adrian, 1941), частота РРП у лягушки может достигать 20 Гц, у кролика (Bartley, Bishop, 1933) около 5 Гц и т. д.
Разрядами последействия ряд исследователей называют и другие ЭЭГ явления — вспышки высокой частоты неизвестной природы. Так, например, Бремер и Бонне (Bremer, Bonnet, 1950) описали так называемый «быстрый разряд последействия» с частотой до 60 Гц, а Эдриан, Джаспер, Айджмон-Марсан и сотрудники (Adrian, 1941; Ajmone-Marsan, Stoll, 1951; Jasper et al., 1952) списали «судорожный разряд последействия», возникающий в структурах мозга после их длительной и сильной стимуляции.
В ряде случаев отмечается, что РРП имеют локализованный характер: возникают в специфическом релейном таламическом ядре и соответствующей ему проекционной зоне коры мозга. При одиночном раздражении неспецифических ядер таламуса также возникают РРП, более широко распространенные по коре.
Вызванные волны
Вызванные волны (ВВ)— явление, видимо, очень близкое, а в ряде случаев, как мы убедимся далее, часто тождественное разряду последействия. Условия их возникновения и локализация, как правило, совпадают с таковыми для РРП. Под ВВ подразумевается процесс провокации в электрокортикограмме (ЭКоГ) тех или иных характерных волн, которые в норме могут характеризовать основной рисунок спонтанной активности мозга. Так, например, во время стадии начальной фазы сна у человека и животных внешнее афферентное раздражение (вспышка света) может спровоцировать появление в ЭКоГ α-волны, являющейся компонентом α-ритма, характерного для ЭЭГ спокойного состояния человека. Та же вспышка может вызвать и целый цуг α-волн (вспышка α-активности), в этом случае говорят о появлении РРП. То же относится и к некоторым другим видам ВВ.
К-комплекс и вертекс-потенциал (Vertex, V-potential)
Видимо, правильнее было бы рассматривать вместе с вторичными ответами коры больших полушарий. Но так как эти ответы описаны под этим названием лишь для ЭЭГ человека, а механизмы их спорны, имеет смысл рассмотреть их отдельно. Кроме того, К-комплекс может возникать и спонтанно.
К-комплекс состоит из двух-трехфазной острой волны с последующим разрядом последействия (σ-послеразряд) (рис. 3). Вначале полагали, что этот ответ является специфическим ответом коры, так как он возникал в ЭЭГ в начальной стадии сна у человека только на звуковое раздражение и регистрировался локально в теменной области полушарий.
Однако дальнейшие работы (Кац, 1959; Пучинская, 1963, 1967, и др.) показали, что К-комплекс возникает при самых разнообразных раздражениях и может наблюдаться не только в начальной стадии сна. Он был отнесен к неспецифическим вызванным потенциалам. К-комплекс возникает в определенной стадии сна и спонтанно, нередко сопровождается вегетативными и двигательными реакциями и не наблюдается во время парадоксального сна (Шаповальников, 1971).
Ряд исследователей придают К-комплексу важное функциональное значение, рассматривая его как электрографическое отражение активности специального антипробуждающего механизма. Есть даже попытка связать К-комплекс со зрительными образами во сне: чем больше проявляется К-комплексов на звук у человека во сне, тем больше и ярче зрительные образы во сне (Fernander, 1969). Некоторые физиологи электрогенез К-комплекса связывают с механизмами, лежащими в основе petit mal эпилепсии.
Вертекс-потенциал впервые описал Девис (Davis, 1939; Devis et al., 1938), затем Гасто (Gastaut, 1951, 1952), Банко (Bancaud et al., 1953) и др. Он регистрируется в области макушки (вертекса) черепа (т, е. там же, где и К-комплекс). При действии различных раздражителей обычно он выражен либо в виде острого пика или медленной однофазной или двухфазной волны (см. рис. 3). Такой нестабильный характер формы ответа (Кац, 1958, 1959; Русинов, 1969, и др.) зависит, видимо, от повторности применяемого раздражения. Так, В. С. Русинов (1969) указывает, что в процессе применения раздражителя (например, звука), прежде чем вертекс-потенциал приобретает форму пика, он проходит стадию низкоамплитудной медленной волны. При дальнейших применениях раздражителя он «растягивается» во времени. Изучая динамику изменения зертекс-потенциала по мере применения раздражителя, отмечая также широкие вариации его формы у разных испытуемых и даже у одного и того же испытуемого во время одного сеанса исследования, Кац приходит к выводу, что и К-комплекс является одной из вариаций этого ответа. С этих позиций К-комплекс и вертекс-потенциал являются вариациями одного и того же ответа — неспецифического ответа коры больших полушарий человека.
Вертекс-потенциал был сравнен со вторичным генерализованным по коре неспецифическим ответом Форбса и Морисона. Затем это мнение было отвергнуто на том основании, что вертекс-потенциал локален. Однако в работах В. С. Русинова с сотрудниками и других исследователей было показано, что вертекс-потенциал является действительно неспецифичным ответом, он возникает на самые различные раздражения, имеет большой латентный период (70—90 мс) и регистрируется не только в области вертекса, но и в других областях коры.
Так, Кац (1959) наблюдал явление распространения ответа на другие области коры больших полушарий, что сопровождалось нарастанием его амплитуды. Данные лаборатории В. С. Русинова, а также данные Ларссона (Larsson, 1960) опровергают высказанную ранее точку зрения Освальда (Oswald, 1959), что неспецифический ответ является артефактом и возникает в результате сдвига кожи на черепе при движении ушей. Неспецифический ответ коры больших полушарий человека может быть использован как ЭЭГ показатель неспецифических влияний на кору больших полушарий. Примером использования, этого ответа служат работы лаборатории В. С. Русинова (Кац. 1959; Пучинская, 1963, 1967; Соколова, 1954; Русинов, 1969, и др.). Что касается ЭЭГ животных, то о неспецифическом ответе, видимо, гомологе неспецифического ответа коры человека, мы будем говорить в специальном разделе нашего курса. Укажем только, что К-комплекс (или вертекс-потенциал) описан и у обезьян (Cohn, 1969; Bert et al., 1969).
Е-волна
Волна «ожидания», или Е-волна, описана Уолтером (Walter, 1963) и названа им «когерентной негативной вариацией» (КНВ). Возможно, она является измененным вертекс-потенциалом в процессе выработки временной связи. Генез ее остается пока неясным. Топография Е-волны ограничена передней и центральной областями головы, включая Фронтальную, моторную и соматосенсорную зону. Е-волна следует сразу за условным сигналом, развивается очень медленно и имеет большую длительность.
Уолтером (1965) она была найдена в следующем опыте. Применялись два раздражителя — звуковой щелчок и ритмический свет — с интервалом в 1 с (рис. 4). Изолированно примененный щелчок вызывает положительную волну (рис. 4, А), замаскированную большим отрицательным компонентом. Ответы на изолированные серии вспышек содержат несколько кратких отрицательных компонентов, также наложенных на небольшое положительное колебание (рис. 4, Б). Когда щелчки применяются вместе со вспышками с интервалом в 1 с (рис. 4, В).
Каждая запись—усредненные ответы на 12 предъявлений раздражителя. Отклонение вверх соответствует электронегативности оба ответа воспроизводятся с небольшими изменениями. После этого испытуемый получает инструкцию нажимать на кнопку в ответ на второй раздражитель (во время вспышек света), что прекращает вспышки, которые являются безусловным подкреплением. В этом случае, в ответ на условный сигнал — щелчок— возникает большая отрицательная волна, которая и была названа Е-волной, или волной «ожидания» (испытуемый ожидает вспышек света, чтобы их включить, нажав на кнопку). Такая картина наблюдается длительное время (рис. 4, Г), пока внимание испытуемого сохраняется, и он точно нажимает на кнопку. Однако, если исключить подкрепление (вспышки света), Е-волна полностью исчезает.
Е-волна возникает сходным образом и в экспериментах, в которых испытуемый должен принять решение в уме, вместо того чтобы выполнять двигательный акт. Необходимо только, чтобы он проявил некоторое умственное усилие в ответ на второй раздражитель (мысленно представил нажатие на кнопку).
Уолтер полагает, что Е-волна имеет корковую природу, так как она инвертирует, если электрод, введенный в мозг человека, проходит через поперечник коры. Это свидетельствует, по его мнению, об ее происхождении в поверхностном плексиформном слое коры. Он также считает, что она отражает деполяризацию сети апикальных дендритов в результате разрядов пирамидных нейронов и не исключает роли гуморальных факторов в ее происхождении.
Некоторые исследователи полагают, что волна «ожидания» имеет место и в ЭЭГ у животных. Так, например, Камп и Ван Риян (Kamp, Rijan, 1970) показали, что при выработке у собаки пищевого двигательного условного рефлекса похожий негативный сдвиг потенциала имеет место в ЭЭГ в период, предшествующий получению пищи.
Основная (фоновая, спонтанная) электрическая активность головного мозга
По внешнему характеру основную активность мозга можно разделить на три вида (см схему):
- Регулярную, или ритмическую, активность, характеризующуюся длинной серией тех или иных волн с незначительной вариацией их частоты;
- Нерегулярную, или аритмическую, активность, состоящую из волн различной длительности л амплитуды;
- Пароксизмальную активность, возникающую на фоне обычной спонтанной активности в виде определенных групп высокоамплитудных волн и комплексов, амплитуда которых внезапно нарастает и спадает.
Однако, прежде чем рассмотреть все эти виды активности, дадим общую характеристику волнам и комплексам, которые либо могут целиком составлять основную активность, либо периодически появляться на ее фоне.
Острая волна. Впервые была описана Джаспером (Jasper, 1941). Спорадические быстрые колебания, с быстрой крутизной нарастания и более пологой крутизной нисходящей части. Длительность волны лежит в пределах 80—200 мс (рис. 5).
Спайк. Спорадические быстрые колебания длительностью около 20 мс с быстрой крутизной нарастания и спада восходящей и нисходящей фазы. При отведении с поверхности мозга длительность спайков может равняться 10 мс. Спайки с длительностью 5—10 мс И. С. Беритов и сотрудники (1943) обозначили как иглоподобные волны. Как острые волны, так и спайки могут возникать внезапно, через нерегулярные интервалы на фоне основной активности. Они могут быть положительные или отрицательные, одиночные и множественные, двухфазные и монофазные, нерегулярные и ритмические (см. рис. 5).
Группы спайков и острых волн определяются как разряды или вспышки. Как правило, спайки и острые волны связаны с патологией, хотя могут иметь место и в обычной ЭЭГ животного и человека.
Комплекс спайк-волна. О нем мы уже говорили ранее. Необходимо только отметить, что могут иметь место вариации этого комплекса, например, две спайк-волны или множественные спайк-волны, острая и медленная волна, когда спайк достигает длительности 80—200 мс, а медленная волна 500—1000 мс.
Дельта (δ)-волны. Длительность δ-волн составляет 250—500 мс. Если они собраны в регулярные серии, то составляют ритм электрической активности мозга — δ-ритм (см. рис. 7).
Комплекс медленных волн. Группа из двух и более медленных волн, повторяющаяся с определенным постоянством формы (см. рис. 5).
Пилообразные волны. Повторяющиеся ритмические острые волны определяют как пилообразную активность, или пилообразные волны (см. рис. 5).
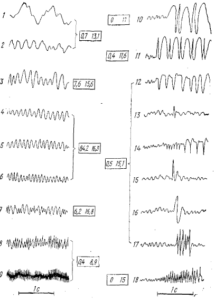
Всем перечисленным выше видам волн и комплексам большое внимание уделяется в клинической ЭЭГ. Спайки, острые и пилообразные волны и комплексы типа спайк-волна объединяют под названием судорожных, или эпилептиформных, потенциалов. На рис. 5 дана классификация электрической активности ЭЭГ человека (Gibbs, 1950). Левая колонка (1—9) показывает различные типы ритмической активности мозга человека, а правая — эпилептическую активность. В пределах цифр от 3 до 7 нормальная ЭЭГ человека и слабые отклонения от нормы (3 и 7). На 1 и 9 крайние степени, на 2 и 8 — значительная степень патологии. ЭЭГ здорового человека почти лишена эпилептических потенциалов (допускается содержание небольшого количества медленных и быстрых волн). Цифры, ограниченные квадратами левого столбца, показывают процент времени, которое тот или иной вид активности может занимать в ЭЭГ взрослого здорового человека; у правого столбца — процент времени, которое занимает данная активность в ЭЭГ больных эпилепсией.
Предложены и другие способы классификации ЭЭГ здорового и больного человека. 1
Характеристика волн, встречающиеся в виде одиночных волн или составляющие ритмы электрической активности нормальной ЭЭГ животных и человека
Тэта-(θ) волны. Их длительность составляет 150— 200 мс. Имеют место как в ЭЭГ человека, так и животных, хотя происхождение их, видимо, различно.
Ламбда-(λ) волны. Одиночные монофазные положительные волны впервые описаны у человека в затылочных областях коры, при том у небольшого процента испытуемых, Гасто и Эвансом (Gastaut, 1951; Evans, 1953). Авторы полагали, что они являются артефактом в результате движения глаз, хотя Грин (Green, 1957) отрицает это и связывает их с механизмом зрения. Длительность λ-волн порядка 300 мс.

Ламбда-волны были описаны в затылочной области коры кошек Родесом и др. (Rodes et al., 1962) и Сторм ван Лёвеном (1965) у собак (рис. 6). Эти исследователи считают, что λ-волны имеют отношение к движению глаз. Предполагается, что они могут служить показателем степени «интереса», с которым животное смотрит. Чем больше количество λ-волн, тем сильнее «интерес».
Альфа-(α) и бета-(β) волны. Длительность α-волн у человека составляет 90—100 мс. Они могут встречаться как в виде отдельных волн, так и составлять регулярный α-ритм. У животных (млекопитающих) в затылочной области коры регистрируются как отдельные альфаподобные полны, так и альфа-подобный ритм (Гусельников, Супин, 1968). Этот вопрос будет рассматриваться нами специально. Что касается β-волн, то они составляют только β-ритм в ЭЭГ человека и у животных не описаны.
Регулярная, ритмическая электрическая активность головного мозга животных и человека (или ритмы электрической активности)
К специфическим ритмам мозга, возникающим спонтанно или при определенных условиях относятся (см. схему): δ-ритм, θ-ритм, α-ритм, β-ритм, роландический, или μ-ритм, σ-ритм (или ритм веретен, веретенообразный ритм), обонятельный ритм, спайк-волна ритм и дыхательный ритм.
Из всех этих ритмов относительно хорошо изучены: α-ритм, θ-ритм, σ-ритм, обонятельный ритм, спайк-волна ритм; элекрогенез других ритмов ЭЭГ изучен слабо.
Ритмы электрической активности складываются из волн, следующих друг за другом с определенным постоянством и имеющих приблизительно одинаковую длительность и форму. Частоты всех ритмов укладываются в диапазон от 1 до 100 кол/с, а амплитуда от 5 до 100— 150 мкВ (последнее относится только к ЭЭГ, регистрируемой с поверхности скальпа или черепа). Ритм, наиболее выраженный в ЭЭГ, в данный момент называют доминантным ритмом.

Дельта-ритм. Впервые дельта (δ)-ритм в ЭЭГ человека был классифицирован Уолтером (Walter, 1936), а впоследствии он был описан и у животных, хотя аналогичность их механизмов еще не ясна. Он состоит из ритмических медленных волн длительностью от 250 до 500 мс или даже до 1000 мс. Отсюда, частота его колеблется от 1 до 4 в 1 с (рис. 7). Некоторые авторы полагают, что δ-ритм— ритмическая активность и с частотой 6—8 кол/с называют ее промежуточной медленной активностью. (табл. 1)
δ-ритм наблюдается в ЭЭГ человека, находящегося под наркозом или при поражении кортикальных отделов мозга (опухоль или острый травматический очаг). Максимальная амплитуда его регистрируется тогда на границе опухоли или очага. Это место и носит название δ-фокус. δ-ритм может встречаться и в ЭЭГ здорового человека как во время сна, так и при бодрствовании. Однако в последнем случае амплитуда его не должна превышать 20—30 мкВ (Жирмунская, 1959). Что касается электрогенеза δ-ритма, то пока точных данных на этот счет нет.
Гутман и Милюкас (1969), Гринявичус и соавторы (1969) рассматривают δ-волны как результат изменения поляризации пирамидных клеточных тел коры. Ройтбак (1969) высказывает предположение об аналогии δ-волн с длительными отрицательными потенциалами, возникающими при сильном прямом раздражении коры и допускает участие нейроглии в их генезе.
Долгое время считалось, что δ-активность, возникающая при самых различных физиологических и патологических состояниях, совершенно одинакова. Однако постепенно появились факты зависимости характера этой активности от порождающих ее условий (Gastaut et al., 1961, и др.).
В пределах δ-активности различают более частые и более медленные компоненты. В ЭЭГ выделяют полиморфные и моноритмичные δ-волны, однако в основу существующих классификаций кладут обычно признак распространенности δ-активности по мозгу; в этом случае она разделяется на локальную и генерализованную (Walter, 1936, 1962; Жирмунская, 1963, и др.). До сих пор, однако, не ясны механизмы широкой генерализации этого ритма, определяющие его возникновение и функциональное значение.
На основании экспериментального изучения δ-активности А. М. Гурвич (1969) предлагает разделить ее на известные уже полифазные δ-волны (ПДВ) с частотой 1—3,5 кол/с, обычно непостоянной формы и комплекс стандартной формы, состоящий из негативных волн длительностью 300—500 мс. Последние волны, где бы они ни возникали, отличаются стандартностью формы и были названы стандартным медленным комплексом (СМК) (рис. 7).
Таблица 1. Ритмы ЭЭГ и частотные характеристики по классификации различных авторов (из Кожевникова и Мещерского, 1963)
| Ритм | Частота, кол/с, по Девис (1938) | Частота, кол/с, по Юнгу (1939) | Частота, кол/с, по Джасперу (1941) | Частота, кол/с, по Гибсу и Гибс (1950) | Частота, кол/с, по Хиллу и Пару (1952) | Частота, кол/с, по Шоубу (1951) | Частота, кол/с, по Страуссу, Остоу и Гринштейну (1952) | Частота, кол/с, по П. И. Гуляеву (1960) | Частота, кол/с, по рекомендации Междунарожной федерации обществ электроэнцефалографии и клинической нейрофзиологии |
|---|---|---|---|---|---|---|---|---|---|
| Дельта (δ) | 0,2—4 | 1—7 | 1—6 | 0,5—5 | 0,5—3,5 | 0,5—3,5 | До 6 | 0,5—3 | Менее 4 |
| Тэта (θ) | — | — | — | — | 4—7 | 4—7 | — | 4—7 | 4—8 |
| Промежуточная медленная активность | 4—8 | — | — | — | — | — | 6—8 | — | — |
| Альфа (α) | 8—13,5 | 8—13 | 8—12 | 6—13 | 8—13 | 8—13 | 8—13 | 8—13 | 8—13 |
| Промежуточная быстрая активность | 13,5—20 | — | — | — | — | 14—17 | 13—15 | — | — |
| Бета (β) | Выше 20 | 14—30 | 18—32 | 14—50 | 14—25 | 18—30 | 15—30 | 14—35 | Выше 13 |
| Очень быстрая активность | — | — | — | — | — | Выше 30 | — | — | — |
| Гамма (γ) | Выше 30 | — | 35—55 | — | Выше 26 | — | — | 35—55 | — |
Тэта(θ)-ритм — складывается из ритмических медленных волн с частотой 4—7 кол/с. Впервые описан и классифицирован как θ-ритм Уолтером и Довейем (Walter, Dovey, 1944). Для ЭЭГ человека принят международный стандарт 4—8 кол/с (рис. 8).
Ряд исследователей полагает, что у человека самостоятельный θ-ритм не существует, и его относят к промежуточной активности между δ- и α-ритмом.
У животных (млекопитающих) θ-ритмом обозначают ритмическую активность с частотой 4—7 кол/с, которая возникает в новой коре, но лучше выражена в лимбических структурах мозга, межуточном мозге и стволе при общей активизации животного, вызванной любыми раздражениями — болевыми, афферентными (при ориентировочной реакции), электрическим раздражением активирующей ретикулярной системы ствола мозга, введением животному антихолинэстеразных препаратов и т. д. Впервые θ-ритм был найден в гиппокамповой коре, где он лучше всего выражен (Gerard el al., 1936; Jung, Kornmiiller, 1938; Green, Arduini, 1953).
Термин θ-ритм (для животных) имеет ряд синонимов: гиппокампальный ритм; гиппокампальный θ-ритм, стресс-ритм и ритм напряжения (Полетаева, Гусельников, 1967). Чаще применяют термин θ-ритм, помня, однако, о том, что аналогичный по частоте ритм ЭЭГ человека мы не можем пока аналогизировать с θ-ритмом животных. θ-ритм описан у многих видов животных. Так, анализ θ-ритма кролика, кошки и обезьяны был проведен Грином и Ардуини (1953). Он описан у ежей, австралийского фалангера, крысы и морской свинки (Liberson, Akert, 1955; Weiss, Fifkova, I960; Adev et al. 1960; Oshima et al., 1962).
У человека выраженность θ-ритма в ЭЭГ зависит от возраста, фона основной активности, степени умственного напряжения. Признаком патологии, видимо, надо считать высокое содержание симметричных θ-волн, а также асимметричные и пароксизмальные θ-волны у взрослого человека в состоянии бодрствования. θ-ритм отражает вовлечение в патологический процесс, видимо, глубоких отделов мозга. Однако Е. А. Жирмунская (1959) полагает, что низкоамплитудный (25—35 мкВ) θ-ритм в ЭЭГ человека может составлять один из компонентов нормальной ЭЭГ. И. С. Берптов и др. (1943) считали, что медленные волны 4—6 кол/с в ЭЭГ здоровых людей являются слиянием 2—3 волн α-ритма; они обозначали их η-волнами. Эта точка зрения по нашла общего признания.
Альфа (α)-ритм состоит из волн частотой около 10 кол/с (от 8 до 13 кол/с) длительностью в среднем 90—120 мс. α-волны имеют «синусоидальную» форму и амплитуду порядка 20—60 мкВ (рис. 9). Могут, однако, наблюдаться высоко- и низкоамплитудные α-волны. Впервые термин α-ритм был введен, как мы уже говорили, Бергером в 1929 г., и ряд исследователей (например, Adrian, Maltews, 1934) назвали этот ритм бергеровским ритмом, α-ритм можно наблюдать во многих областях мозга человека. Он регистрируется как в обширных областях коры, так и во многих подкорковых образованиях— таламусе, базальных ядрах и др. (Okuma et al., 1954; Sem-Jacobsen ci al., 1955, 1956). Во всех точках коры мозга a-активность имеет одну и ту же частоту и постоянные фазовые соотношения.
α-ритм характерен для ЭЭГ взрослого здорового человека при состоянии бодрствования и закрытых глазах. Любая активация, усиление внимания и т. д. ведут к угнетению, блокаде α-ритма. При наркозе и естественном сне α-ритм также исчезает. Таким образом, α-ритм является своеобразной характеристикой определенного функционального состояния коры больших полушарий. Сдвиг в сторону от оптимального уровня — уровня α- активности (понижение или повышение степени активации) ведет к угнетению α-активности (Кратки, 1957). Если на фоне бодрого, спокойного состояния (когда в ЭЭГ хорошо выражен α-ритм) давать активирующие воздействия, наблюдается блокада α-ритма и те же воздействия на фоне дремотного состояния (когда α-ритм отсутствует) вызывают вспышку α-ритма. Частота α-ритма у разных испытуемых заметно отличается, по она крайне стабильна для каждого испытуемого (Брейзье. 1962), Обычно амплитуда α-волн испытывает определенную модуляцию: периодически то увеличивается, то ослабляется (сквик-эффект, по Уолтеру). В результате этого α-волны собираются в своеобразные веретена (см. рис. 9), длительность которых колеблется от 0,5 до 5 с. и более. В ряде случаев такое веретено α-ритма начинается с более высокочастотных волн, но через 0,5—1 с частота их снижается до обычной. Амплитуда α-ритма в разных точках может быть неодинакова.
Ранее α-ритм связывали преимущественно со зрительным анализатором, так как именно в затылочной области коры он имеет большую амплитуду и значительно легче блокируется световыми раздражениями. Предполагали, что именно здесь расположен первичный корковый очаг α-ритма, однако это не подтвердилось при более тщательных исследованиях.
Было показано (Garouttc et al., 1956, 1959), что могут быть различные случаи распределения α -ритма . В одних случаях изучение фазовых соотношений α-колебаний показало, что возможно постепенное волнообразное распространение α-волн от задних областей коры к передним, α-колебания в затылочной области коры возникали на 12—20 мс раньше, чем в центральных, и на 40—50 мс раньше, чем во фронтальных. Однако можно наблюдать и противоположный эффект — более раннее появление α-ритма в передних областях и почти так же часто, как и в затылочной области; причем α-волна из передней области может распространяться к затылку. Близкие данные получили и другие исследователи (Ю. В. Дубикайтис и В. В. Дубикайтис, 1962, 1963; Cooper, Mandy-Castle, 1960, и Др.). Это даст основание полагать, что α-ритм может первично возникать в разных зонах коры мозга человека, хотя фокус его активности привязан, как правило, к затылочной области коры. Последнее подтверждается исследованиями электрической активности мозга человека при нарушении зрения, которые показали прямую связь α-ритма со зрительной функцией.
Впервые Эдриан (Adrian, Mattews, 1934; Adrian, Jamagiva, 1935) высказал предположение о роли зрительной афферентации в формировании α-ритма. До последнего времени существовали разноречивые данные по этому вопросу. Данные, полученные в клинике человека, свидетельствовали об исчезновении α-ритма при различных повреждениях зрительного анализатора, вызывающих слепоту или значительную редукцию функции зрения. Ряд других исследователей полагают, что α-ритм прямо не зависит от зрительной афферентации и что он имеет место даже у слепых.
Л. А. Новикова (1966), подвергнув этот вопрос специальному экспериментальному анализу, пришла к заключению, что у слепых людей и особенно у слепорожденных α-ритм не регистрируется у людей с дефектом зрения по мере возрастания остроты зрения степень выраженности α-ритма на ЭЭГ увеличивается. Предпринятый Л. А. Новиковой анализ механизмов этого явления привел к заключению, что депрессивный характер ЭЭГ слепых и в том числе отсутствие α-ритма связано со снижением тонуса коры и обусловлено выключением ведущей в жизни человека зрительной афферентации.
В настоящее время различают α-активность и α-ритм. Первая обычно состоит из одиночных α-волн с длительностью от 80 до 125 мс. Эта α-активность регистрируется в любых мозговых структурах; α-ритм определяется как регулярная волновая активность с частотой порядка 10 Гц и регистрируется преимущественно в теменно-затылочных областях.
Из работ ряда авторов известно, что у человека к 13 годам электрическая активность мозга приобретает черты, свойственные ЭЭГ взрослых людей. ЭЭГ детей 10—12-летнего возраста по степени выраженности α-ритма в затылочных областях коры отличается от ЭЭГ взрослых людей незначительно, α-ритм остается четко выраженным до 50—60 лет. Однако в электроэнцефалографии детского возраста существуют до сих пор противоречия по вопросу характеристики отдельных компонентов ЭЭГ и их классификации (Фарбер, 1969). Ряд авторов полагают, что α-ритм формируется у ребенка из ритмических колебаний 3—4 в 1 с, преобладающих в ЭЭГ маленьких детей. Уолтер (Walter, 1953) и другие авторы считают, что увеличение частоты α-ритма с возрастом не происходит. Они полагают, что в процессе развития сменяются нейронные механизмы генерации ритмов, разных по частоте и своему генезу.
Многие авторы относят к α-ритму маленьких детей ритмы ЭЭГ 3—5 в 1 с, другие — 6—8 в 1 с, а третьи — 8—9 в 1 с. Другие категорически возражают против этого, полагая, что все эти колебания являются не генетическим аналогом «будущего» α-ритма, а представляют собой самостоятельные колебания другой природы.
Очевидно, что выявление удобной модели α-ритма У животных может сыграть важную роль для изучения его генеза (Гусельников, Супин, 1968). Активность, очень сходная с α-ритмом, наблюдается у обезьян (Cavennes, 1962), кошек (Clark, Ward, 1945; Крагин, 1962) и других животных. Описан и еще один вид α-подобной активности у кролика, которая, возможно, аналогична α-ритму человека. Это ритмические колебания потенциала в затылочной коре бодрствующего спокойного кролика, имеющие частоты 4—5 в 1 с (Bartley, 1940; Bishop, Clare, 1952, и др.). Эта активность имеет ряд свойств, сближающих ее с α-ритмом человека: фокус ее также локализован в затылочной коре в области проекции зрительного анализатора; этот ритм угнетается, как и α-ритм человека, при общей активности животного и при действии световых раздражителей. Все это дает основание считать, что он может являться аналогом α-ритма (Bartley, 1940).
Роландический ритм был открыт в 1952 г. Гасто и затем описан в работах других авторов (Gastaut et al., 1957; Klass, Bicford, 1957; Роже, Воронин, Соколов, 1958; Chatrian et al., 1959, и др.). Роландический ритм еще называют μ-ритмом, гребенчатым, дугообразным или аркообразным ритмом.
Этот ритм регистрируется с черепа или с обнаженной поверхности мозга только у человека з области роландовой борозды, где всегда находится его фокус. Он напоминает по своей частоте и форме α-ритм, но отличается отпето по ряду внешних признаков и по физиологическим свойствам. Его частота 7—11 кол/с. Форма веретенообразная, но часто может иметь вид однофазных колебаний характерной формы, отчего за ним в литературе закрепилось название «аркообразный», или «wicket»-ритм.
Роландический ритм в норме можно зарегистрировать лишь у небольшого числа испытуемых (например, у 15 из 500, т. е. в 3% случаев, Chatrian et al., 1959). Он отличается от α-ритма по ряду функциональных свойств. Если α-ритм наиболее чувствителен к световым раздражениям, то роландический ритм — к тактильным и проприоцептивным раздражениям, а также к воображаемым движениям. Депрессия ритма при всех этих раздражениях лучше всего выражена в противоположном полушарии, что говорит об особой роли проприоцептивной афферентации в его блокаде. Л. А. Новикова отмечает, что у слепых на общем фоне депрессии корковой ритмики происходит формирование роландического ритма, она связывает это с компенсаторным усилением деятельности двигательного анализатора.
Роландический ритм, вероятно, может быть идентифицирован с рядом описанные ранее вариантов α-ритма (Кожевников, Мещерский, 1963), прецентральным α-ритмом (Jasper, Andrews, 1938), высокоамплитудным α-ритмом (Schutz, Muller, 1951) и α-подобной активностью (Maddocks et al., 1951). Иногда его определяют как β-вариант-ритм, подчеркивая его функциональную и тонографическую общность с β-ритмом, что вряд ли оправдано.
Несмотря на различия роландического ритма и α-ритма, есть принципиальное сходство между ними: так же как и α-ритм, он наблюдается при неактивном, расслабленном состоянии двигательной системы, прямо зависит (блокируется) от активации ведущего для него двигательного анализатора, имеет близкую частоту с α-ритмом и т. д. Можно ожидать, что они действительно являются различными модификациями одного и того же процесса. Именно в связи с этим можно отметить, что при отведении от височных областей черепа человека у 50% испытуемых умственные нагрузки вызывают не депрессию, а появление в ЭЭГ ритмической веретенообразной активности с частотой (8—12 кол/с) и амплитудой 20—30 мкВ (Kennedy et al., 1948). Авторы обозначили волны этого ритма каппа (κ)-волны, а ритм — κ-ритм (или Кеннеди волны). Таким образом, и в этой области коры полушарий наблюдается активность, сходная с α-ритмом. Возможно, что все эти сходства являются отражением еще не ясных общих механизмов и общих принципов генеза всех этих ритмических колебаний коры, получающих лишь «специфическую окраску», в основе которой лежит та или иная модальность афферентного потока. Что касается гомологов роландического ритма у животных, то некоторые исследователи считают таковым «сенсомоторный» ритм в коре у кошки (Roth et ah, 1967; Chase, Harper, 1971; Howe, Sterman, 1972; Rougeul et al., 1972).
Бета (β)-ритм коры человека состоит из быстрых волн длительностью до 40—50 мс и частотой 14—30 кол/с (см. рис. 9). Впервые термин β-ритм был введен Бергером. Обычно β-ритм связывают с сенсомоторной системой.
И. С. Беритов (1948) полагает, что этот ритм коры определяется деятельностью нервных кругов и цепочек коры, что наблюдается обычно при активном состоянии ретикулярной формации ствола мозга; при ее блокаде β-ритм исчезает.
У большинства животных в разных отделах мозга, в том числе и в коре больших полушарий, можно зарегистрировать сходные и более высокочастотные ритмы. Ряд исследователей их обычно не классифицируют как β-ритм, оставляя этот термин лишь для ЭЭГ человека, тем более что об общности их генеза ничего не известно.
Однако И. С. Беритов (1969) относит к β-активности высокочастотные ритмы в ЭЭГ кошки (до 500 в 1 с) и в ЭЭГ обезьян (до 200—300 в 1 с). У человека к β-активности он относит волны с частотой от 250 до 480 в 1 с. Чем выше частота β-ритма, тем ниже его амплитуда.
β-ритм человека регистрируется, как правило, в лобных отделах мозга. Генерализованная по коре низкоамплитудная быстрая β-активность является показателем патологии. Так, Флюгель (Fliigel, 1968) отмечает, что в группе больных с высокочастотными β-волнами (больше 30 в 1 с) отмечен большой процент психовегетативных синдромов.
Оценивая функциональное значение β-ритма на разных этапах развития патологического процесса при эпилепсии, опухоли мозга и травме, И. В. Введенская и др. (1971) рассматривают его как показатель включения или усиления структур мозга, участвующих в компенсаторных процессах. Ими показано, что наличие β-ритма в ЭЭГ больных в восстановительном периоде после травмы черепа и мозга имеет благоприятное прогностическое значение,
Гамма (γ)-ритм состоит из волн низкой амплитуды не более 15 мкВ, имеющих частоту 30—35 кол/с. Он выражен лучше в передних отделах мозга. Если его амплитуда выше 15 мкВ, то ЭЭГ рассматривается как патологическая (Кожевников, Мещерский, 1963).
Сигма (σ)-ритм впервые для человека был описан Лумисом с соавторами (Loomis et al., 1935) при исследовании электроэнцефалографического выражения различных стадий сна.

σ-ритм является одним из основных и наиболее четко выраженных компонентов спонтанной ЭЭГ млекопитающих, находящихся в состоянии естественного сна или сна подобных состояниях, вызванных некоторыми фармакологическими и нейрохирургическими вмешательствами. Большинство исследователей считают, что σ-ритм появляется в ЭЭГ человека при наступлении легкого сна (Gibbs, 1950; Dement, Kleitman, 1957; Hess, 1964, и др.). Спонтанный σ-ритм представляет собой вспышки ритмических электрических колебаний с частотой до 10—16 в в 1 с. Появление в виде отдельных групп волн с постепенно нарастающей и затем убывающей амплитудой (рис. 10) является характерным внешним признаком σ-ритма, за что он и получил другие названия — веретенообразная активность (spindle activity), взрывная или вспышечная активность (burst activity), веретенообразные вспышки (spindle burst), α-подобный ритм животных, сонные веретена (sleep spindles), барбитуровые веретена (barbiturate spindles) и ряд других. Надо полагать, что термины веретена и вспышки должны быть сохранены за целым классом ЭЭГ колебаний, имеющих характерные амплитудные модуляции (например, за α-ритмом, роландическим ритмом, обонятельным ритмом) и обозначение этим термином α-ритма— не целесообразно. Термин α-подобные также неприемлем, т. к. α-ритм и α-ритм — совершенно различные виды электрической активности мозга (Гусельников, Мухаметов, 1967, 1969). Название сонные веретена и барбитуровые веретена удобно сохранить за разновидностями этого ритма. Мы и дальше, будем все его модификации обозначать σ-ритмом. Спонтанные его проявления будем называть σ-веретенами, а вызванные одиночным раздражением мозга — σ-послеразрядами. За некоторыми разновидностями σ-ритма, видимо, рационально сохранить наиболее привычное обозначение. К спонтанному σ-ритму относятся σ-веретена естественного сна (сонные веретена), наркотического сна. вызванного барбитуратами (барбитуровые веретена), препаратов мозга с различными хирургическими нарушениями активирующих систем и при некоторых других состояниях мозга.
Вызванный σ-ритм может быть получен либо одиночным возбуждением механизма этого ритма (σ-послеразряды), либо ритмическим возбуждением на резонансной частоте 10—14 в 1 с.
К σ-послеразрядам относятся специфические σ-послеразряды, которые возникают на таламо корковом уровне специфических афферентных систем в ответ на короткий адекватный стимул или одиночное электрическое раздражение любого уровня данной системы (рис. 10). Многие авторы (Dempsey, Morison, 1943; Chang, 1950, Bremer, 1958, Andersen, Eccles, 1962) называют эти ритмические колебания по-разному.
Под специфическими послеразрядами мы будем подразумевать послеразряды, возникающие только в слуховой и соматической афферентных системах. Кроме того, σ-послеразряды могут быть вызваны коротким электрическим раздражением неспецифического таламуса, неокортекса, хвостатого ядра (веретена хвостатого ядра — caudate spindles), мезенцефалической ретикулярной формации и некоторых других структур (см. рис. 10). Необходимо отметить, что при ритмическом возбуждении механизма σ-ритма возникает реакция вовлечения, если раздражается неспецифический таламус при связанные с ним структуры, или реакция усиления, если раздражается специфический таламус. В соответствии с этим целый ряд спонтанных и вызванных ритмических процессов, часто изучавшихся изолированно, имеет, как мы убедимся, по существу, одни и тот же механизм и представляет собой модификацию одного и того же ритма.
Обонятельный ритм. Под обонятельным ритмом подразумевается ритмическая электрическая активность обонятельного анализатора животных и человека, впервые описанная у животных Эдрианом (Adrian, 1942, 1956) и названная им обонятельными вызванными волнами.
Обонятельные вызванные волны в обонятельной луковице (ОЛ) и ряде других образовании мозга (в основном, структурах обонятельного мозга) регистрируются у человека и позвоночных животных всех классов (рис. 11 и табл. 2), В отличие от млекопитающих и человека, у рыб, амфибий, рептилий и птиц они являются, видимо, единственным видом королю выраженной ритмической электрической активности мозга (Гусельников, 1965; Гусельникова, Гусельников, 1975). Вероятно, незначительные изменения структурных особенностей обонятельной луковицы и ее связей в ряду позвоночных свидетельствуют о том, что вызванные волны имеют одинаковую природу у представителей различных классов и видов позвоночных. Такое предположение в некоторой степени подтверждается сходством электрических реакции обонятельной луковицы у различных классов позвоночных, а также отсутствием заметных их различий у макро- и микросматиков (Oltoson, 1963).
Таблица 2. Частота вызванных волн в обонятельной луковице позвоночных
| Объект исследования | Частота волн, кол/с | Авторы |
|---|---|---|
| Рыбы | 6—8 | Гусельникова, Гусельников, 1975 |
| Амфибии | 6—9 | Geets, 1948; Takagi, Shibuya 1960 |
| Рептилии | 15—25 | Winkel, Caspers, 1953; Гусельников, 1956 |
| Птицы | 15—25 | Гусельников, 1964 |
| Млекопитающие | ||
| Насекомоядные (еж) | 15—45 | Adrian, 1942 |
| Грызуны (кролик) | 55 | Гедеванишвили, 1955 |
| Кошка | 45 | Adrian, 1942 |
| Собака | 20—40 | Ueki, Domino, 1961 |
| Обезьяны | 40 | Hughes, Mazurowski, 1962; Ueki, Domino, 1961 |
| Человек | 38—70 | Гедеванишвили, 1955; Sem — Jacobsen et al., 1956 |
Вспышка вызванных обонятельных волн имеет форму веретена и обычно появляется, в фазе вдоха, хотя их можно наблюдать и в фазе выдоха (Ueki, Domino, 1961). Обонятельные вызванные волны можно получить в остром опыте у всех видов животных «протягиванием» воздуха через носовую полость трахеотомированного животного (Новикова, Хволес, 1953; Гусельников, 1964; Adrian, 1942, и др.). Отсюда обонятельный ритм и получил название «обонятельные вызванные волны», хотя он регистрируется спонтанно во время вдоха или выдоха животного или особенно во время ориентировочной реакции «принюхивания».
У представителей различных классов позвоночных частота волн во вспышке разная. В ряду рыбы — амфибии—рептилии—млекопитающие наблюдается явная тенденция к увеличению частоты. Что касается ритмической обонятельной активности, регистрируемой в других структурах мозга, с которыми обонятельная луковица имеет прямые связи, то было показано, что у кролика (Новикова, Хвотес, 1953) обонятельные вызванные волны могут иметь место на всем пути oт обонятельной луковицы вплоть до переднего гипоталамуса. У кошек эти волны имеют место в амигдало-препириформной области и гиппокампе (Gault, Coustau, 1965), у обезьян и собак в обонятельной полоске, пириформной и препириформной коре, медиальной миндалине., заднем гипоталамусе (Domino, Ueki, 1960), у крысы — в препириформной коре (Гусельников, 1964), у ящерицы (Winkel, Caspers, 1953) и черепахи (Гусельников, 1956, 1964) в гиппокампальной коре и у птиц в septum, в медиальном гиперстриатуме (Гусельников, 1964) и т. д.
Таким образом, почти у всех представителей различных классов позвоночных в центральных структурах обонятельного анализатора возникают обонятельные волны, сходные по конфигурации и частоте с вызванными волнами обонятельной луковицы.
Спайк-волна ритм. Характеристики этого ритма мы уже касались ранее, при рассмотрении комплекса спайк-волна.
Дыхательный ритм. При регистрации ЭЭГ могут наблюдаться артефакты, связанные с ритмом дыхания. Обычно это связано с механическим смещением отводящих электродов на коже черепа в момент вдоха и выдоха.
Однако в ЭЭГ имеет место и физиологический дыхательный ритм — S-ритм (от слова «spiro» — дышу) (рис. 12). Этот ритм не является артефактом, но изучен пока крайне недостаточно. Обычно он регистрируется при специальных условиях (Ливанов, Поляков, 1945; Бут, Климова-Черкасова, 1969, и др.). Считается, что S-ритм в ЭЭГ проявляется при ухудшении функционального состояния коры (Григорьев, 1958; Гурвич, 1966; Бут, Климова-Черкасова, 1969, и др.). Так, Гурвич (1966) показал, что этот ритм регистрируется особенно четко при умирании организма.
Однако определенная зависимость синхронизации ЭЭГ здорового человека от дыхательного цикла была найдена рядом исследователей (Roth, Klingberg, 1970; Faber et a]., 1970, и др.).
Еще в 1936 г. А. И. Смирновым, а позднее М. В. Сергиевским (1950) и др. было показано влияние дыхательного центра на кору больших полушарий. В 1954 г. Ройтбак получил прямые доказательства иррадиации импульсов из дыхательного центра на кору больших полушарий. Он обнаружил, что судорожные потенциалы в отравленном стрихнином участке коры наступают точно по ритму дыхания. В наших экспериментах с использованием той же методики (Гусельников, 1960) был показан универсальный характер влияния дыхательного центра на вышележащие отделы мозга у позвоночных разного уровня эволюционного развития (см. рис. 12).
Источником S-ритма в ЭЭГ являются каудальные отделы ствола мозга, и большинство исследователей связывают его с дыхательным центром (Григорьев, 1958; Королькова, 1966; Латаш, 1959; Бут и Климова-Черкасова, 1969, и др.). Бут и Климова-Черкасова полагают, что S-ритм связан с распространением к коре импульсации по неспецифическим афферентным путям из ретикулярной формации каудального отдела ствола мозга через внутреннюю капсулу (экстраталамически).
Надо полагать, что восходящее тоническое влияние дыхательного центра связано с облегчающим действием на корковые нейроны (Угрюмов, 1947; Sharpless, Jasper, 1956) и имеет, видимо, постоянный характер, но проявляется в ЭЭГ при разных сдвигах функционального состояния мозга, в том числе и при ориентировочной реакции (Ливанов, Поляков, 1915; Бут и Климова-Черкасова, 1969). Специальный анализ связи ЭЭГ с дыхательным ритмом провел А. А. Генкин (1971). Он пришел к выводу не только о наличии такой связи, но и предполагает наличие локальных влияний дыхательного центра на определенные области коры. Тонические влияния дыхательного центра имеют место не только на уровне коры, но и на уровне, спинного мозга (Орбели, Кунстман, 1961; Латаш, 1959), а также таламуса и гипоталамуса (Бут, Климова-Черкасова, 1969).
Нерегулярная, или аритмическая, фоновая активность головного мозга
Обычно она состоит из волн самой различной длительности и амплитуды. Могут быть разные, случаи проявления этой активности. Так, у многих животных как в коре полушарий, так и в других отделах мозга нерегулярная, аритмичная активность может составлять основной рисунок ЭЭГ и уступать место ритмической, регулярной активности лишь при определенных физиологических состояниях (стресс-состояние, ориентировочная реакция, сон и т. д.).
В других случаях эта активность может наблюдаться в «интервалах» между различными «вспышками» той или иной регулярней активности. Провести какие-либо градации этой активности сложно, тем более что она, по сути дела, является суммой самых различных колебаний и в каждом отдельном случае необходим специальный анализ составляющих ее компонентов.
Пароксизмальная активность
Включает четко ограниченные группы высокоамплитудных волн, которые обычно возникают и резко выделяются на фоне более низкоамплитудного основного ритма (Кожевников, Мещерский, 1963). Так, пароксизмальные вспышки ритмов с частотой 10 и 25 кол/с могут наблюдаться в нормальной ЭЭГ при состоянии бодрствования, а вспышки 14 кол/с — при состоянии сна (вспышки σ-веретен) (рис. 13). У больных эпилепсией могут появляться самые различные пароксизмальные ритмы — комплексы спайк-волн, вспышки медленных волн и т. д. Пароксизмальные, ритмы, характеризующиеся фиксированной частотой, возникают внезапно, прогрессивно нарастают до максимума и внезапно прекращаются.
Продолжая дальше классификацию электрических процессов мозга, необходимо рассмотрев изменения фоновой ритмики (см. схему).
Изменения фоновой активности структур головного мозга животных и человека “возникают “при самых различных условиях: действии внешних раздражителей, естественных состояниях сна и бодрствования, ориентировочной и условнорефлекторной деятельности, течении сложных безусловных реакций, раздражении мозговых структур, действии различных физических и гуморальных факторов и т. д. Ряд этих воздействий вызывает общие сдвиги текущей спонтанной активности мозга, часто не равнозначные в различных мозговых структурах.
Поэтому, чтобы в дальнейшем легче было разобраться в генезе и динамике изменений фоновой активности, отметим несколько групп общего типа реакций, имеющих место в коре больших полушарий животных и человека:
- Реакцию активации;
- Реакцию инактивации;
- Искусственные способы изменения фоновой ритмики.
Реакция активации. Еще Бергер (Berger, 1929) отметил, что открывание глаз, внешние раздражения, умственное напряжение приводит к депрессии, угнетению, блокаде α-ритма. В 1937 г. Рейнбергер и Джаспер (Reinberger, Jasper), исследуя ЭЭГ и поведение котики, наблюдали пробуждение животного при действии разных афферентных раздражений. Они выяснили, что все применяемые раздражения, вызывающие пробуждение и бодрствование, сопровождались сменой в ЭЭГ медленных ритмов на низковольтную быструю активность. Эта реакция была названа ими реакцией активации. Они же отметили генерализованный характер этой реакции. Позже Моруцци и Мэгун показали, что прямое раздражение ретикулярной формации ствола мозга воспроизводит все изменений, характерные для активации, связанной с бодрствованием, ничем не отличающейся от реакции при действии внешних раздражителей. С этого момента реакцию угнетения медленной ритмики, или реакцию активации, начинают чаще называть реакцией пробуждения, или эраузал-реакцией (arousal reaction).(см. рис. 14)
Часто применяется и другой термин для этой реакции— десинхронизация. Этот термин не точен и не применим для всех случаев реакции активации. В относительной степени он может быть использован в реакции, когда четкая ритмическая (синхронизированная) активность мозга сменяется высокочастотной активностью. Мы будем пользоваться термином реакция активации.
У человека (Соколов, 1962) в зависимости от исходного фона активация с ЭЭГ проявляется: а) в усилении быстрых колебаний на фоне уже развившейся депрессии α-ритма, б) постепенной депрессией α-ритма и увеличением доли в суммарной ЭЭГ β- и γ-колебаний. в) депрессией медленных δ и θ-колебаний и сменой их или высокоамплитудным α-ритмом, или же повышением β-активности.
Реакции активации противопоставляют реакцию инактивации (Соколов, 1962), которая связана со снижением уровня возбуждения в коре. Эта реакция, в зависимости от исходного уровня активности, выражается в появлении α-ритма вместо быстрой низкоамплитудной активности, появлении медленных δ и θ-волн вместо β- и α-колебаний, дальнейшем замедлении и возрастании амплитуды медленных волн, уже присутствующих в фоновой активности.
Изменение ритмики электрических колебаний коры, как в случае реакции активации, так и при инактивации, может быть различной длительности и носить фазический и тонический характер (Соколов, 1956). По данным Шарплесса и Джаспера (Sharpless, Jasper, 1956), тоническая реакция связана с участием стволовой части ретикулярной системы, а фазическая — с неспецифической системой таламуса.
Обычно реакция активации сопровождает ориентировочную деятельность человека и животных, и она, как мы указывали, как правило, связана с генерализованным по коре угнетением спонтанной ритмики и появлением быстрой, высокочастотной активности. Однако у ряда животных ориентировочная реакция сопровождается появлением синхронизированных ритмов в ЭЭГ. Так, у кроликов и крыс в ответ на афферентные раздражения (звук, свет и т. д.) возникают синхронизированные колебания с частотой 4—7 в 1 с (Green, Arduini, 1954; Соколова, 1954; Новикова, Фарбер, 1959; Семпохина, 1958; Гусельникова, 1958; Чжоу-Шао-Цы, 1958; Банцекипа, 1962, и др.). Особенно четко эти синхронизированные ритмы при ориентированной реакции наблюдаются в древних структурах мозга — гиппокампе, гипоталамусе и ретикулярной формации.
Систематические исследования, проведенные нами (Гусельников, 1964) на рыбах, амфибиях, рептилиях и птицах, показали, что у хронически оперированных животных с вживленными электродами ориентировочная реакция сопровождается в электрограмме различных отделов мозга сложным характером изменении суммарной активности, который зависит от многих факторов, и прежде всего от исходного фона активности, который в свою очередь определяется внешними условиями. В связи с этим реакция может выражаться либо в угнетении, либо в усилении фоновой активности, что, видимо, связано со спецификой развития в филогенезе функций ретикулярной формации ствола мозга.
Искусственные способы изменения фоновой ритмики
Включают изменения активности мозговых структур, вызванные прямым действием ряда химических и физических факторов: локальная аппликация на поверхность мозга различных фармакологических препаратов, прямое электрическое раздражение, анодная и катодная поляризация нервных структур, прямое действие механических и физических факторов и т. п. Все эти воздействия входят в арсенал средств экспериментальной нейрофизиологии, используемых для анализа механизмов электрогенеза суммарной активности мозга. Также воздействия могут вызвать изменения фоновой активности, не имеющие места в естественных условиях. Так, например, при аллокации кусочка фильтровальной бумаги, смоченной 2—2,5%-ным КСl, на поверхность мозга возникает вначале локальное, а затем распространяющееся по коре полное подавление электрической активности. Это явление получило название по имени автора — распространяющаяся депрессия Лео (Leao, 1944). Распространяющаяся депрессия как прием исследования была широко использована для решения ряда интересных вопросов нейрофизиологии коры. Этот и ряд других приемов искусственного воздействия на фоновую активность мы встретим в разных разделах нашего курса.
Длительные электрические (сверхмедленные) колебания потенциала
Наличие медленных длительных потенциалов в коре головного мозга было показано многими авторами (Русинов, 1947, 1962; Швец, 1958; Аладжалова, 1960, 1962; Brookhart et al., 1958, и многие другие). Значительно сложнее и запутанней обстоит дело с классификацией всех этих медленных и сверхмедленных колебаний потенциала. Эти потенциалы называют: апериодические медленные колебания, медленные длительные потенциалы, изменения уровня постоянного потенциала, сверхмедленные колебания, гладкие потенциалы, квази- постоянный потенциал и т. д.
В 1969 г. Н. А. Аладжаловой была предложена классификация длительных потенциалов, которой мы будем придерживаться. По характеру проявления она предложила разделить их на:
- Постоянные (стабильные) потенциалы;
- Апериодические сдвиги постоянного потенциала;
- Ритмические (сверхмедленные) колебания постоянного потенциала. По временному параметру последние делятся на секундные, минутные, многоминутные и часовые.
В целом колебательные потенциалы с такими временными периодами следует определять как сверхмедленные в отличие от δ и θ-ритма ЭЭГ. К числу длительных потенциалов головного мозга следует отнести и потенциалы, регистрируемые в коре больших полушарий при прямом электрическом ее раздражении.
Трудности, связанные с классификацией длительных потенциалов головного мозга, определяются прежде всего тем, что их происхождение остается до сих пор не совсем ясным; имеется много гипотез на этот счет, но ни одна из них, как отметил Ройтбак (1969), не является еще фактически достаточно обоснованней. Остается еще далеко неясным и вопрос о функциональном значении этих потенциалов, хотя определенная связь, например сдвига постоянного потенциала с изменениями ряда других суммарных реакции мозга, говорит об определенной их роли в интегративных функциях нервной системы.
Постоянные (стабильные) потенциалы регистрируются при отведении с разных участков мозга. Между поверхностью коры и белым веществом, а также между поверхностью коры и спинномозговой жидкостью и кровяным руслом имеет место постоянная разность потенциалов. При естественном сие и при наркозе потенциал коры смещается в положительную сторону, при бодрствовании— в отрицательную.
Апериодические сдвиги постоянного потенциала наступают при искусственных раздражениях различных нервных структур, в том числе и коры, адекватной или электрической стимуляции рецепторов, воздействиях на метаболизм мозга, при безусловной и условнорефлекторной деятельности, изменений кровообращения и аппликации различных веществ. Эти сдвиги постоянного потенциала (или, как еще говорят, сдвиги уровня постоянного потенциала, Русинов, 1969) могут носить локальный или диффузный характер, что зависит, видимо, от условий опыта (применяемых электродов и способов отведения, уровня наркоза, функционального состояния животного и т, д.), а также от характера применяемой стимуляции.
Сверхмедленные ритмические колебания потенциала — колебания, близкие по форме к синусоидальным, с частотой 6—8 в 1 мин (при амплитуде 0,3—0,8 мВ) — или секундный ритм (рис. 15, А) и колебания с частотой 0,5— 2 в 1 мин (при амплитуде 0,5—1,5 мВ) —минутный ритм (рис. 15, Б). В разных образованиях головного мозга эти колебания проявляются различно. Наблюдаются они преимущественно в коре больших полушарий и в гипоталамусе. В ретикулярной формации ствола и таламуса они не найдены.
Длительные потенциалы коры на ее прямое электрическое раздражение
В ответ на относительно сильный стимул, нанесенный на поверхность коры, после начального отрицательного потенциала длительностью 20— 30 мс, возникает медленный отрицательный потенциал (МОП), продолжительность которого равна сотням миллисекунд и даже секунд (Ройтбак, 1965). МОП возникает при напряжении стимула в десять раз большем, чем пороговый стимул для вызова дендритного потенциала (Ройтбак, 1969). Он достигает амплитуды 2—3 мВ и продолжительности 0,5—3 с; латентный период равен 15—20 мс. Регистрируется до 3 мм от пункта раздражения только в верхних слоях коры (рис. 16). При ритмических электрических раздражениях коры возникает постоянное отрицательное отклонение, которое, как считает Ройтбак, является результатом суммации медленных отрицательных потенциалов, МОП используется в экспериментальной ЭЭГ для выяснения некоторых особенностей коркового электрогенеза.
Footnotes
- Кожевиков В. А., Мещерский Р. М. Современные методы анализа электроэнцефалограммы. М., 1963.
Характеристики альфа-ритма электроэнцефалограммы у больных бронхиальной астмой
Для цитирования: Мельникова Т.С., Краснов В.Н., Мартынова Н.В. Характеристики альфа-ритма электроэнцефалограммы у больных бронхиальной астмой. Доктор.Ру. 2019. № 1 (156). С. 41–45. DOI: 10.31550/1727-2378-2019-156-1-41-45
08 февраля 2019
Цель исследования: провести сравнительный анализ параметров основного ритма электроэнцефалограммы (ЭЭГ) человека — альфа-ритма — у больных бронхиальной астмой (БА) и здоровых людей.
Дизайн: открытое контролируемое исследование.
Материалы и методы. В исследовании участвовали 57 пациентов с частично контролируемой БА легкой и средней тяжести и 50 здоровых испытуемых. У больных БА отмечались умеренные расстройства аффективного спектра (13–17 баллов по шкале депрессии Гамильтона).
Результаты. При спектральном анализе ЭЭГ у больных БА выявлены диффузное снижение спектральной мощности альфа-ритма при усилении медленноволновой активности, преимущественно тета-спектра, и ослабление функциональных связей между всеми областями коры, особенно между симметричными височными областями.
Заключение. Полученные данные указывают на особую мозаику пространственно-временных соотношений функционирования различных отделов коры и межкорковых взаимодействий при БА. Это позволит дифференцированно подходить к оценке функциональной активности различных корковых зон и их участия в интегративной деятельности мозга. Данные показатели могут служить дополнительными критериями при диагностике и выборе терапевтических подходов у больных БА.
Авторы заявляют об отсутствии возможных конфликтов интересов.
Краснов Валерий Николаевич — д. м. н., профессор, руководитель отдела клинико-патогенетических исследований МНИИП — филиала ФГБУ «НМИЦ ПН им. В.П. Сербского» Минздрава России. 107076, г. Москва, ул. Потешная, д. 3. eLIBRARY.RU SPIN: 9644-6970. E-mail: [email protected]
Мартынова Наталья Владимировна — научный сотрудник отдела клинико-патогенетических исследований МНИИП — филиала ФГБУ «НМИЦ ПН им. В.П. Сербского» Минздрава России. 107076, г. Москва, ул. Потешная, д. 3. eLIBRARY.RU SPIN: 1318-6671. E-mail: [email protected]
Мельникова Татьяна Сергеевна — д. б. н., главный научный сотрудник лаборатории патологии мозга МНИИП — филиала ФГБУ «НМИЦ ПН им. В.П. Сербского» Минздрава России. 107076, г. Москва, ул. Потешная, д. 3. eLIBRARY.RU SPIN: 6584-2792. E-mail: [email protected]
Приблизительно 300 млн человек в мире имеют диагноз бронхиальной астмы (БА) [1]. При этом заболеваемость продолжает возрастать, и к 2025 году ожидается увеличение популяции больных БА еще на 100 млн человек [1, 2]. Распространенность в разных возрастных группах, прогрессирующее повышение заболеваемости и затрат на лечение приводят к увеличению бремени этой болезни как для бюджета здравоохранения, так и для общества и самих пациентов, что делает БА значительной медицинской и социальной проблемой [1-3]. При современном уровне доказательной медицины системный подход к проблеме БА возможен только на основе междисциплинарных исследований на стыке различных областей знания — пульмонологии, психиатрии, физиологии, психофизиологии, нейропсихологии, социологии. В этом направлении нужен определенный научный и практический прорыв, который позволит нам приблизиться к разработке необходимых при БА методов персонализированной медицины. БА многие годы трактовалась как одна из классических психосоматических болезней. Не углубляясь в историю вопроса о трансформации самого понятия «психосоматические болезни» и прежних трактовок в этом аспекте БА, можно констатировать, что, согласно современным представлениям, БА является самостоятельной нозологической формой. БА — хроническое воспалительное заболевание дыхательных путей, характеризующееся обратимой бронхиальной обструкцией и гиперреактивностью бронхов. Возникновение и прогрессирование БА может служить примером сложных психосоматических и соматопсихических взаимоотношений и взаимосвязей [4]. В подавляющем большинстве случаев на всех этапах развития БА в той или иной мере отмечается наличие психопатологических расстройств аффективного спектра, чаще тревожно-депрессивного комплекса. Степень выраженности психопатологических включений в основную клиническую картину заболевания создает дополнительные трудности в лечении больных БА. Одним из подходов к объективной оценке дезорганизации функционального состояния корковой мозаики ЦНС является анализ электрической активности мозга, особенно основного ритма ЭЭГ — альфа-активности [5, 6]. Многочисленные исследования показали, что развитие депрессивных состояний сопровождается нарушениями структуры всех частотных диапазонов ЭЭГ, но в большей степени изменения касаются основного ритма ЭЭГ — альфа-ритма [7-10]. Альфа-ритм — ритмические колебания с частотой 8-13 Гц и средней амплитудой 30-70 мкВ, регистрируется преимущественно в затылочных областях при закрытых глазах в состоянии спокойного бодрствования и максимально возможном расслаблении мышц. Ритм блокируется световой стимуляцией, концентрацией внимания и умственными нагрузками. Альфа-диапазон при депрессиях может значительно усиливаться или редуцироваться, меняется пространственное распределение ритма [8, 11-13]. По современным представлениям, генерация альфа-ритма связана с реверберацией импульсной активности по интеркортикальным и таламо-кортикальными нейронным сетям, а выраженность его обусловливает синхронизацию функциональной активности различных мозговых систем, в частности определяет связь получаемой от афферентной системы организма информации с механизмами оперативной памяти, регулируя адаптационные процессы. Поэтому альфа-диапазон ЭЭГ традиционно привлекает повышенное внимание исследователей благодаря его высокой чувствительности к разнообразным внешним воздействиям и тонким изменениям функционального состояния коры мозга, сопровождающим сенсорные, моторные, когнитивные и мнестические процессы [14, 15]. Характер изменений альфа-активности при депрессивных расстройствах зависит в первую очередь от особенностей депрессивного синдрома. Так, повышение индекса альфа-ритма свойственно больным с «большой депрессией», а его снижение в десинхронизированном типе ЭЭГ выявляется при дистимических расстройствах, т. е. при депрессиях невротического уровня [16-18]. Рядом авторов предложена нейрофизиологическая модель депрессии, постулирующая изменение фронтальной асимметрии — снижение левополушарной активности [19-25]. Исследователи связывают особенности изменения альфа-активности с доминированием тоскливого, тревожного или апатического аффекта, подтверждая тем самым общепринятое положение о связи структуры ЭЭГ с функциональной активностью корковых зон. Наблюдаемые при этом особенности профиля вегетативной нервной системы позволяют предположить, что одним из механизмов перестройки корковой ритмики является дисфункция диэнцефальных подкорковых структур [26]. Цель исследования: провести сравнительный анализ параметров основного ритма ЭЭГ человека — альфа-ритма — у больных БА и здоровых людей.
МАТЕРИАЛЫ И МЕТОДЫ В исследовании при условии добровольного информированного согласия участвовали 57 пациентов (38 женщин и 19 мужчин) с частично контролируемой БА легкой и средней тяжести и с расстройствами аффективного спектра. Диагноз БА устанавливался в соответствии с международными согласительными документами (Global Initiative for Asthma, 2017). Возраст пациентов варьировал от 18 до 65 лет (средний возраст — 53,7 ± 10,9 года). Группу контроля составили 50 практически здоровых лиц, сопоставимых по возрасту и полу. Проводилось комплексное клинико-инструментальное обследование: функциональное исследование легких (спирометрия, функция внешнего дыхания), ЭЭГ. Для оценки и анализа психопатологических расстройств аффективного спектра использовалась клинико-психопатологическая карта, разработанная В.Н. Красновым [27]; шкалы Гамильтона для оценки тревоги и депрессии; тест-контроль БА. Общей психопатологической характеристикой обследованных больных БА была гипотимия с тревожным компонентом, при этом прослеживалась тенденция трансформации тревоги в тревожно-тоскливое состояние. Количество баллов по шкале депрессии Гамильтона составило 13-17. Литературные данные свидетельствуют о различных профилях нейрофизиологических показателей у правшей и левшей, поэтому по опроснику латеральных признаков [28] в группы испытуемых отбирались правши. ЭЭГ регистрировали с помощью аппаратно-программного комплекса для топографического картирования электрической активности мозга «Нейро-КМ» (Россия) с полосой пропускания от 0,1 до 45 Гц. Запись ЭЭГ осуществляли монополярно от симметричных лобных (F3, F4), центральных (C3, C4), теменных (P3, P4), затылочных (О1, O2), передне-височных (F7, F8), средне-височных (T3, T4) и задне-височных (Т5, Т6) корковых зон (схема 10-20%, четные каналы отведения — от корковых зон правого полушария, нечетные — левого). Референсным электродом служили объединенные ушные клипсы. Характеристики и топографическое распределение ритмов ЭЭГ определяли с помощью анализа ЭЭГ методом быстрого преобразования Фурье с усреднением не менее 30 эпох по 2 секунды с последующим картированием по системе BRAINSYS (Россия). Проводили спектральный и когерентный анализы ЭЭГ. Величину когерентности (КОГ) вычисляли между всеми 14 отведениями корковых областей в диапазоне 7-13 Гц, а также с шагом 1 Гц. Определяли и величину средней когерентности (СрКОГ) каждой области со всеми остальными корковыми зонами — показателя функциональной активности выбранной зоны в интегральной деятельности мозга. При межгрупповом сравнительном анализе спектральной мощности (СМ) ритмов использовали приведение показателей к нормализованному типу через логарифмирование (LnCM), а сравнение параметров КОГ проводилось по формуле:
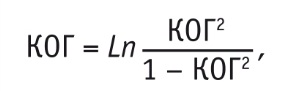
где КОГ 2 — квадрат модуля КОГ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ Данные визуального и спектрального анализов электроэнцефалограммы Наиболее выраженные изменения фоновой ЭЭГ обследованных больных наблюдались в виде двух вариантов: низкоамплитудной дизритмии и усиления синхронизации в медленноволновых диапазонах. В первом случае отмечались снижение амплитуды биоэлектрической активности, редукция альфа-ритма, сглаженность регионарных различий по альфа-индексу, усиление диффузной низкоамплитудной медленноволновой активности. При втором варианте имели место билатеральные вспышки альфа- и тета-волн, возникающие синхронно во всех областях и указывающие на явления ирритации на уровне стволовых структур мозга. Характерной особенностью альфа-ритма стала слабая выраженность или полное отсутствие модуляции в веретёна. Независимо от типа изменений ЭЭГ-картирование групповых спектральных характеристик ЭЭГ выявляло нарушения топики основного ритма — зональные различия по альфа-индексу были слабо выражены. Несмотря на то что в группу исследования включались только правши, фокус альфа-ритма у обследованных пациентов был нечетко обозначен в правой затылочной области, что характерно для нормы, а в отдельных случаях доминировал в затылочной области левого полушария, что указывало на более значительное снижение функциональной активности левого полушария. При спектральном анализе ЭЭГ абсолютная СМ альфа-ритма у больных была ниже, чем у испытуемых контрольной группы, во всех отведениях ЭЭГ. Диффузное снижение СМ альфа-ритма приводило к тому, что при сопоставлении с остальными ритмами ЭЭГ СМ альфа-ритма не лидировала, а занимала позицию после СМ медленноволновых ритмов, чаще после тета-диапазона. Изменение соотношений отдельных ритмов ЭЭГ у больных БА особенно наглядно можно проследить при анализе %СМ — составляющей доли в процентах альфа-ритма от совокупности СМ всех ритмов ЭЭГ в данной области коры. На рисунке 1 показана разность %СМ между показателями у больных и испытуемых контрольной группы. Во всех корковых зонах снижение %СМ альфа-ритма составляло более 12%. Наиболее значительная редукция (около 20%) — в затылочных областях с акцентом в правом полушарии, что и отражалось в нивелировании зональных различий корковых зон по выраженности альфа-индекса. Рис. 1. Отличия %спектральной мощности альфа-ритма у больных бронхиальной астмой от нормы (p < 0,05 при модуле 1,7)
У больных отмечено смещение модального значения СМ ведущего ритма в более низкий частотный диапазон — 8-9 Гц (против 10 Гц у здоровых испытуемых), что отражает переход корковой ритмики на функционирование в более энергосберегающих режимах ( рис. 2 ). Следующими по значению СМ в альфа-диапазоне являлись полосы 9-10 и 10-11 Гц, а самыми низкими — высокочастотные полосы 11-12 и 12-13 Гц. Средний показатель СМ по всем корковым зонам (СрСМ) высокочастотных составляющих альфа-ритма в 4 раза ниже СрСМ доминирующей частоты 8-9 Гц.
Рис. 2. Структура альфа-ритма — средние значения 1 Гц полос спектральной мощности (СМ) по всем отведениям
По сравнению с показателями контрольной группы ( рис. 3 ) у больных БА полоса 8-9 Гц была снижена в меньшей степени, чем остальные. Наибольшая редукция полосы 10-11 Гц отмечалась в затылочных областях с небольшим акцентом в правом полушарии. В этих корковых зонах полосы 9-10, 11-12 и 12-13 Гц были уменьшены почти в равной мере. Во фронтальных корковых зонах закономерность иная — в большей степени редуцирована полоса 9-10 Гц, затем следуют полосы 10-11, 11-12 и 12-13 Гц. Эти данные указывают на значительную деформацию структуры основного ритма у больных. Генераторы альфа-ритма у пациентов работают в особом режиме, значительно отличающемся от нормы. Рис. 3. Изменение структуры альфа-ритма у больных бронхиальной астмой по сравнению с нормой во фронтальных (F3 слева и F4 справа) и затылочных (О1 слева и О2 справа) областях коры (p < 0,05 при модуле 1,7)
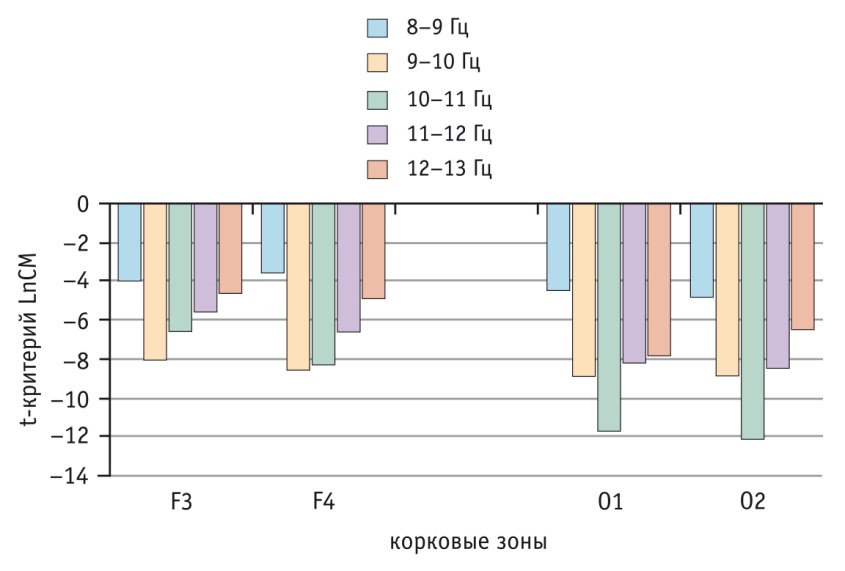
Данные когерентного анализа электроэнцефалограммы Наиболее низкие значения КОГ у больных БА выявлены в диапазонах длиннодистантных связей. Так, лобно-затылочные величины КОГ (О1-F3, О1-F4, О2-F4 и О2-F3) колебались в диапазоне 0,05-0,08. Короткодистантные связи имели существенно более высокие значения КОГ: затылочные области с теменными зонами (О1-Р3, О2-Р4) — 0,34 и 0,48, фронтальные области с близлежащими областями (F3-С3, F4-С4) — 0,75 и 0,74. Величина СрКОГ по альфа-диапазону у больных БА по сравнению с таковой у участников контрольной группы ( рис. 4 ) по всем корковым зонам была значительно снижена (р < 0,05). Рис. 4. Сравнение величины средней когерентности (СрКОГ) по альфа-диапазону у больных и испытуемых контрольной группы
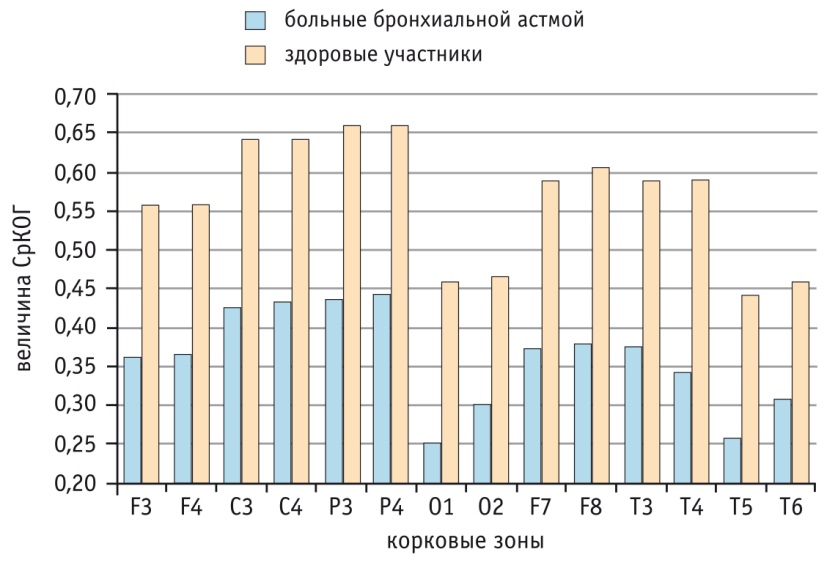
Во фронтальных, центральных и теменных зонах обоих полушарий уменьшение достигло полуторакратного размера, а в височных зонах — несколько меньшего. Самое большое различие СрКОГ по альфа-диапазону между нормой и больными выявлено в затылочных и задне-височных зонах, в частности в левой затылочной области — СрКОГ по альфа-диапазону у больных была почти в 2 раза ниже, чем у здоровых. Полученные данные свидетельствуют о значительном диффузном ослаблении функциональных связей между всеми областями коры. Это касается и межполушарных связей, они уменьшены между всеми симметричными корковыми зонами по основному ритму, хотя и в разной степени. Так, между фронтальными зонами отличия от нормы менее выражены, чем между другими парами корковых областей. Особенно значительное уменьшение межполушарного взаимодействия отмечается между передне-, средне- и задне-височными зонами. Роль отдельных частотных полос альфа-ритма в связях между корковыми зонами неодинакова — основную роль играет низкочастотная полоса 8-9 Гц. Значения СрКОГ этой полосы наибольшие во фронтальных, теменно-центральных зонах, а также в височных зонах правого полушария. Следовательно, у больных БА перестройка структуры основного ритма в более замедленные режимы изменяет и снижает функциональную активность корковых зон. Проведенное клинико-нейрофизиологическое исследование подтверждает наличие сопутствующих БА умеренных тревожно-депрессивных расстройств с тенденцией формирования тоскливого варианта аффективного спектра, на что может указывать усиление медленноволновой активности, преимущественно тета-диапазона.
ЗАКЛЮЧЕНИЕ Исследование альфа-ритма ЭЭГ с применением спектрального и когерентного анализа позволило выявить у больных бронхиальной астмой (БА) особую мозаику пространственно-временных соотношений работы различных отделов коры и межкорковых взаимодействий. Это позволяет дифференцированно подходить к оценке функциональной активности различных корковых зон и их участия в интегративной деятельности мозга на основании спектральных и когерентных показателей альфа-диапазона. Данные показатели могут служить дополнительными характеристиками при диагностике и выборе терапевтических подходов у больных БА.
ЭЭГ-мониторинг или лекция для тех кому любопытно
Электроэнцефалографический мониторинг (ЭГГ -мониторинг) — сегодня это основной метод исследования, цель которого регистрация пароксизмального события с целью проведения дифференциального диагноза между эпилептическими и неэпилептическими состояниями, например, такими как парасомнии, синкопальные состояния, стереотипии и другие.
Широкое распространение метода ЭЭГ-мониторинга в первой половине 90-х гг. произвело буквально революционные изменения в диагностике эпилепсии, позволило распознавать сложные в клиническом отношении судорожные припадки. Стало ясно, что многие виды судорожных приступов не укладываются в классическую картину, описанную в медицинских учебниках, что привело к пересмотру взглядов на диагностику и тактику лечения пациентов.
Проведение ЭЭГ назначается для получения ответов на следующие вопросы:
- Какова природа судорожных приступов — эпилептическая или неэпилептическая? (какую болезнь лечить)
- Какая форма эпилепсии? (как правильно лечить, какие препараты)
- Какая локализация приступа? (постановка вопроса о целесообразности хирургического лечения при неэффективности медикаментозного)
- Как протекает лечение? (изменять , отменять препараты)
Прежде чем приступить к ответу на эти вопросы необходимо понять происхождение этого метода, а потом прийти к плодам, результатам многолетних исследований, которые в изобилии разрослись на этом мощном стволе.
2. Понятия, определяющие ВЭЭГ
2.1. ЭЭГ и эпилепсия
Основные понятия, которые можно выделить, разбирая понятие видео-ЭЭГ-мониторинг — ЭЭГ и эпилепсия.
Вспомним определение эпилепсии: эпилепсия является одним из распространенных хронических заболеваний головного мозга. Согласно определению, эпилепсия — это хроническое заболевание головного мозга, характеризующееся повторными непровоцируемыми приступами нарушения двигательных, чувствительных, вегетативных, мыслительных или психических функций, возникающих вследствие чрезмерных нейронных разрядов (ILAE , 1989).
Электроэнцефалография — метод исследования биоэлектрической активности головного мозга, основанный на определении разности электрических потенциалов, генерируемых нейронами в процессе их жизнедеятельности. Регистрирующие электроды располагают так чтобы на записи были представлены все основные отделы мозга. Получаемая запись — ЭЭГ — суммарная электрическая активность миллионов нейронов, представленной преимущественно потенциалами дендритов и тел нервных клеток: возбудительными и тормозными постсинаптическими потенциалами и частично — потенциалами дендритов и тел нервных клеток. То есть ЭЭГ — как бы визуализированный результат функциональной активности головного мозга.
Здесь наверное стоило бы уделить долю внимания анатомии нейрона и его физиологии.
Нейрон является главной клеткой центральной нервной системы. Формы нейронов чрезвычайно многообразны, но основные части неизменны у всех типов нейронов: тело и многочисленных разветвленных отростков. У каждого нейрона есть два типа отростков: аксон, по которому возбуждение передается от нейрона к другому нейрону, и многочисленные дендриты (от греч. дерево), на которых заканчиваются синапсами (от греч. контакт) аксоны от других нейронов. Нейрон проводит возбуждение только от дендрита к аксону.
Основным свойством нейрона является способность возбуждаться (генерировать электрический импульс) и передавать (проводить ) это возбуждение к другим нейронам, мышечным, железистым и другим клеткам.
Нейроны разных отделов мозга выполняют очень разнообразную работу, и в соответствии с этим форма нейронов из разных частей головного мозга также многообразна.
Многолетние исследования в области нейрофизиологии привели к выводам, что следующие электрические события присущи нейронам и могут вносить в вклад в суммарную биоэлектрическую активность мозга (ЭЭГ ): постсинаптические возбуждающие и тормозные потенциалы (ВПСП , ТПСП), и распространяющиеся потенциалы действия (ПД ). ВПСП и ТПСП возникают либо в дендритах, либо на теле нейрона. ПД генерируются в зоне аксонного «холмика » и далее распространяются по аксону.
Нейрон. Возбуждающие и тормозные ПСП, потенциал действия.
Обычная спонтанная ЭЭГ, ее основные ритмы возникают в результате пространственной и временной суммации постсинаптических потенциалов (ПСП ) большого количества корковых нейронов. Временные характеристики процесса суммации достаточны медленны по сравнению с длительностью ПД.
Определенная степень синхронизации задается различными подкорковыми структурами, которые выполняют роль «водителя ритма» или пейсмекера. Среди них наиболее значимую роль в генерации ритмов ЭЭГ играет таламус.
Потенциалы действия корковых нейронов не играют существенной роли в генерации основных ритмов ЭЭГ, поскольку являются очень короткими. Определяющая роль ПД в формировании паттернов ЭЭГ возникает в ситуациях, когда значительное количество нейронов синхронизируется и одновременно выдает пачки или «вспышки » ПД. Такой режим характерен для пароксизмальных событий, например, для эпилептических приступов и тогда морфология волн ЭЭГ в существенной степени определяется потенциалами действия. При этом острые компоненты ЭЭГ (спайки , острые волны) отражают не отдельные ПД, а скорее всю «пачку » потенциалов действия. Именно таким образом формируются многие эпилептиформные паттерны ЭЭГ, наиболее известным из которых является комплекс спайк-волна. Следует отметить, что подобная модель применима также в объяснении генеза физиологических острых компонентов ЭЭГ.
Таким образом, в генерации ЭЭГ принимают участие и постсинаптические потенциалы и потенциалы действия. Основная ритмика ЭЭГ определяется градуальными изменениями постсинаптических потенциалов благодаря пространственной и временной суммации отдельных ПСП в больших популяциях нейронов, которые относительно синхронизированы и находятся под воздействием подкоркового водителя ритма. Пароксизмальные же события, синхронизирующие значительное количество нейронов, которые продуцируют вспышки потенциалов действия, отвечают за формирование многих эпилептиформных феноменов ЭЭГ, в частности комплексов спайк-волна.
Собственно изучением всего этого процесса и занимается электроэнцефалография.
2.2. История изучения ЭЭГ
Начало изучению электрических процессов мозга было положено Д. Реймоном (Du Bois Reymond) в 1849 году, который показал, что мозг, также как нерв и мышца, обладает электрогенными свойствами.
Начало электроэнцефалографическим исследованиям положил В.В.Правдич-Неминский, опубликовав 1913 году первую электроэнцефалограмму записанную с мозга собаки. В своих исследованиях он использовал струнный гальванометр. Так же Правдич-Неминский вводит термин электроцереброграмма.
Первая запись ЭЭГ человека получена австрийским психиатром Гансом Бергером в 1928 году. Он же предложил запись биотоков мозга называть «электроэнцефалограмма ».
По мере совершенствования компьютерной техники в 1996 году осуществлена методика амбулаторной полиграфической записи посредством 17-канального электроэнцефалографа (16 каналов ЭЭГ и 1 канал ЭКГ) с использованием переносного персонального компьютера (ноутбука ) [Ebersole J. S. et al, 1996].
И наконец к концу 20 века у в арсенале эпилетологических и нейрофизиологических служб получается несколько видов методики ЭЭГ: рутинная ЭЭГ, холтеровское ЭЭГ и ВЭЭГ.
ЭЭГ стал «лезвием бритвы», наиболее качественным и информативным методом диагностики формы эпилепсии и позволяет регистрировать клинико-электроэнцефалографический коррелят эпилептического приступа, что дает возможность установить более точный диагноз и назначить рациональную схему антиэпилептической терапии.
3. Проведение ЭЭГ. Взгляд с 3 позиций
ЭЭГ мониторинг можно рассмотреть с 3 позиций: пациента, врача который проводит исследование в данный момент и со стороны врача, который расшифровывает видео-ЭЭГ-мониторинг постфактум.
Как предисловие стоит просто назвать показания и противопоказания к проведению этого исследования (Авакян )
- Диагностика эпилепсий и эпилептических синдромов.
- Пароксизмальные состояния неясного генеза, вызывающие подозрение на наличие эпилепсии.
- Фармакорезистентные приступы (с целью выявления псевдоэпилептических пароксизмов или уточнения формы эпилепсии).
- Контроль эффективности лечения.
- Медикаментозная ремиссия (объективная констатация ремиссии).
- Подготовка к отмене антиконвульсантной терапии.
- Прехирургическое обследование.
- Субклиническая эпилептическая активность.
- Прогрессирующие когнитивные и поведенческие расстройства у детей.
- Первый судорожный приступ.
Противопоказания:
Противопоказаний к проведению ЭЭГ нет.
3.1. Устройство ВЭЭГ-лаборатории
Сутью метода ВЭЭГ является непрерывная запись ЭЭГ сигнала и видеоизображения пациента в течение длительного времени. Минимальная продолжительность исследования 15 минут, максимальная не лимитирована (до 7-14 суток). Обязательным условием является идеальная синхронизация видеоизображения и ЭЭГ во времени.
Основой системы ЭЭГ-мониторинга является многоканальный усилитель сигналов, имеющий возможность записи 19-32-64-128-канальной ЭЭГ, ЭКГ-канала, датчика дыхания, электромиографических и электроокулографических каналов.
К усилителю подключаются соответствующие регистрирующие датчики. Крепление ЭЭГ-электродов для длительной регистрации производится с помощью специальной шапочки или клеящей пасты. Конструкция электродной системы позволяет пациенту перемещаться по палате, не доставляет неудобств и делает проведение исследования комфортным.
Сигналы с усилителя посредством проводной или беспроводной связи направляются на рабочую компьютерную станцию.
Видеоизображение записывается посредством цифровых видеокамер, количество их может быть произвольным, в большинстве систем предусмотрена возможность использования 1-2 камер.
Обработка результатов производится изучением синхронного изображения ЭЭГ и видеоизображения, скорость прокрутки изображения выбирается произвольно. Программа обработки ЭЭГ включает возможности спектрального и когерентного анализа, программ трехмерной локализации диполя, и других вариантов компьютерного анализа.
Отделение видео-ЭЭГ-мониторинга должно включать в себя 3 основных звена:
- палата пациента, оборудованная видеокамерами, микрофоном, кнопкой пациента для регистрации событий;
- помещение для записывающих станций и персонала, осуществляющего слежение и контроль за пациентом.
- помещение для врачей (ординаторская ), где располагаются станции просмотра и анализа записанных данных. Важным требованием к ВЭЭГ оборудованию является возможность просмотра и обработки ранее записанных исследований или текущего, не прерывая текущее исследование.
3.2. Схемы монтажа, подготовка пациента и начало исследования
Врач, проводящий исследование накладывает электроды либо встроенные в шапочку либо поочередно наклеивает каждый электрод соответственно своему месту. В обычной практике ЭЭГ-регистрирующие электроды располагают согласно международной системе «10 -20».
В соответствии с системой «10 –20» у испытуемого делают три измерения черепа:
- продольный размер черепа — измеряют расстояние по черепу между точкой перехода лобной кости в переносицу (назион ) и затылочным бугром;
- поперечный размер черепа — измеряют расстояние по черепу через макушку (вертекс ) между наружными слуховыми проходами обоих ушей;
- длину окружности головы, измеренной по этим же точкам.
Электроды, расположенные по средней линии, отмечаются индексом Z; отведения на левой половине головы имеют нечетные индексы, на правой — четные.
Отведения в системе «10 –20»:
- фронтальные (F1 , F2., F3 F4, Fz);
- лобные полюса (Fp1 , Fp2);
- центральные (С1 , С2, С3,С4, Сz);
- париетальные (Р1 , Р2 Р3 Р4, Рz);
- темпоральные (T1 , Т2, Т3, Т4, Т5, Тz);
- окципитальные (О1 ,О2, 0z).
Крепление и наложение электродов осуществляется в следующем порядке:
- Электроды подсоединяются к усилителю. Для этого штеккеры электродов вставляются в электродные гнезда усилителя.
- Ватным тампоном, смоченным в спирте обезжириваются места, предназначенные для постановки электродов.
- Непосредственно перед постановкой каждого электрода электродный гель наносится на контактирующую с кожным покровом поверхность. Необходимо помнить, что гель, применяемый в качестве проводника, должен быть предназначен для электродиагностики.
- На пациента надевается шлем/шапка со встроенными поверхностными электродами или каждый поврхностный электрод по отдельности с креплением его при помощи специального клея — коллодия. От практики игольчатых электродов ныне отходят согласно последним исследованиям ученых США и Великобритании. Места расположения электродов определяются соответственно системе расположения электродов. Необходимо помнить, что наложенные электроды не должны вызывать у пациента неприятные ощущения.
- Соответственно обозначениям, указанным на панели усилителя, устанавливаются электроды на предусмотренных системой местах, парные электроды располагаются симметрично.
После правильной установки и калибровки начинается само ВЭЭГ-исследование. В сегодняшней практике используются ВЭЭГ исследования длиной 4-5 часов (утренние /дневные/вечерние), длиной 9-10 часов (ночные ), длиной 24 часа и более (холтеровские ВЭЭГ-мониторинги). Наиболее распространенными сегодня, являются короткие ВЭЭГ-исследования (60 %), далее ночные — 36%, холтеровские — 4-5%
Премедикация перед исследованием, как правило, не проводится, поскольку введение препаратов, не входящих в схему лечения, может менять картину ЭЭГ, что не позволит оценить истинные параметры биоэлектрической активности мозга.
Не рекомендуется отмена или изменение схемы лечения в день исследования, что не позволит оценить эффекты терапии.
Принципиальное значение в диагностике эпилепсии имеет ЭЭГ сна. По мнению ведущих специалистов».. регистрация ЭЭГ в течение одной минуты поверхностного сна дает больше информации для диагностики эпилепсии, чем час исследования в состоянии бодрствования».
4. Понятия нормы и патологии в ВЭЭГ
4.1. Стандарты ВЭЭГ
Альфа ритм. Ритм частотой 8-13 Гц средней амплитуды 50 мкВ (15 -100 мкВ), максимально выражен в задних (затылочных ) отведениях при закрытых глазах. Возможно появление модуляций альфа ритма ( «веретен »), заключающихся в периодическом нарастании и снижении амплитуды волн. Выраженность альфа ритма зависит от многих условий, что необходимо учитывать при анализе ЭЭГ. Представленность альфа ритма на ЭЭГ и его регулярность уменьшаются при открывании глаз, записи в состоянии беспокойства, при активной умственной деятельности (решение задач), а также во время сна. У женщин во время менструации может быть увеличение его частоты. Установлено, что у здорового взрослого человека частота альфа ритма довольно стабильна и является генетически детерминированной.
Мю (роландический , аркообразный) ритм. Ритм аркообразной формы, частоты альфа (обычно , 8-10 Гц). По амплитуде не превышает альфа ритм (обычно , несколько ниже); регистрируется в центральных отделах у 20% здоровых взрослых. У детей данный ритм начинает хорошо определяться с 3-месячного возраста, лучше у девочек. Он не реагирует на открывание глаз, но блокируется с одной стороны при выполнении движений в контралатеральной конечности. Имеет малое диагностическое значение, даже при значительном его усилении или выраженной асимметрии.
Бета ритм. Ритм частотой более 13 Гц, средней амплитуды 10 мкВ; максимально выражен в передних отделах. Типичная частота бета ритма в норме составляет 18-25 Гц, менее распространен ритм частоты 14-17 Гц и крайне редко — свыше 30 Гц, У 70% здоровых людей амплитуда бета ритма не превышает 10 мкВ; и лишь у 3% — превышает 20мкВ. Бета ритм имеет максимальную выраженность в лобно-центральных отведениях. Бета активность несколько усиливается в период сонливости, при засыпании (I стадия сна), во время ФБС и иногда при пробуждении. В период глубокого сна (III , IV стадии фазы медленного сна) амплитуда и выраженность ее существенно снижаются.
Стойкое увеличение амплитуды бета активности свыше 25 мкВ, особенно с распространением её на задние отведения — признак патологии, однако, нозологически не специфичной. Традиционно усиление бета активности ( «excessive fast») связывалось с текущим эпилептическим процессом.
Тета ритм. Ритм частотой 4-7 Гц, по амплитуде, обычно превышающий основную активность фоновой записи. Встречается различной степени выраженности на ЭЭГ у всех здоровых детей. Тета активность начинает регистрироваться в центральных отделах уже с 3-недельного возраста, постепенно нарастая с возрастом и достигая максимума в 4-6 лет. В этом возрасте тета ритм является доминирующим на ЭЭГ у детей. Большинство исследователей считают, что у подростков и молодых взрослых при бодрствовании с закрытыми глазами низкоамплитудная тета активность (не превышающая амплитуды фона) частоты 6-7 Гц с бифронтальным преобладанием является нормальной, если она не превышает 35% фоновой записи.
4.2. Запись во сне
Сон является мощным активатором эпилептиформной активности. Неврологу, а тем более эпилептологу, важно уметь идентифицировать фазы и стадии сна. Известно, что эпилептиформная активность отмечается преимущественно в I и II стадии медленного сна, тогда как во время «дельта сна» и в периоде ФБС она чаще всего подавляется.
В настоящее время для дифференциации стадий сна применяется классификация Dement & Kleitman в модификаций Recbtshaffen & Kales (1968 ), Согласно этой классификации выделяются 2 фазы сна: фаза медленного сна (ФМС ) и фаза быстрого сна (ФБС ),
ФМС (в англоязычной литературе — non-REM sleep) развивается на фоне ослабления влияния активизирующей коры, восходящей ретикулярной формации и усиления активности синхронизирующих тормозящих структур.
В ФМС выделяют 4 стадии.
I стадия сна (дремота ) характеризуется умеренным замедлением основной активности на ЭЭГ. Проявляется постепенным исчезновением альфа ритма и появлением ритмичной тета активности в центральной и лобно-центральной области, Может появляться периодическая ритмическая высокоамплитудная медленная активность частотой 4-6 Гц в лобных отведениях. Длительность I стадии сна у здорового человека составляет не более 10-15 минут.
II стадия сна (стадия «сонных веретен»). Наблюдаются следующие феномены. 1. Характерный признак II стадии сна — появление «сонных веретен» или сигма-ритма. Этот феномен представляет собой ритмические веретенообразно нарастающие и снижающиеся по амплитуде вспышки с частотой 12-16 Гц и амплитудой 20-40 мкВ, преимущественно в центрально-париетальных отделах. Длительность «сонных веретен» колеблется от 0 до 2 сек. Высокоамплитудные и продолжительные (около 3 сек) сонные веретена с преобладанием в лобных отведениях — обычно признак патологии.
III стадия сна характеризуется нарастанием амплитуды и количества медленных волн, преимущественно, дельта диапазона. Регистрируются К-комплексы и «сонные веретена». Дельта волны на эпохе анализа ЭЭГ занимают до 50% записи. Отмечается снижение индекса бета активности.
IV стадия сна характеризуется исчезновением «сонных веретен» и К-комплексов, появлением высокоамплитудных (не менее 50 мкВ> дельта волн, которые на эпохе анализа ЭЭГ составляют более 50% записи. III и IV стадии сна являются наиболее глубоким сном. Они объединены под общим названием «дельта сон».
В фазе быстрого сна (парадоксальный сон, REM-сон) отмечается ослабление влияния тормозящей ретикулярной формации и усиление десинхронизирующих активизирующих механизмов. При входе в ФБС нарастает бета активность. Данная фаза сна характеризуется появлением на ЭЭГ картины десинхронизации в виде нерегулярной активности с одиночными низкоамплитудными тета волнами, редкими группами замедленного альфа ритма и острыми «пилообразными » волнами. ФБС сопровождается быстрыми движениями глазных яблок и диффузным снижением мышечного тонуса. Именно в эту фазу сна у здоровых людей происходят сновидения. Начало сна с фазы быстрого сна или возникновение ее менее, чем через 15 минут после засыпания — признак патологии.
Нормальный сон взрослых и детей состоит из чередования серии циклов ФМС и ФБС. ФМС наиболее выражена в первую половину ночи и занимает 75% всего сна. Во второй половине ночи наиболее представлена ФБС (фаза сновидений), которая занимает около 25% ночного сна. Продолжительность одного цикла сна у детей младшего возраста составляет 45-55 минут; у взрослых 75-100 минут. За ночь у здорового человека возникает от 4 до 6 циклов сна.
4.3. Медленноволновая активность.
Медленноволновая активность. Этот термин включает в себя активность на ЭЭГ в форме замедления ритма по сравнению с возрастной нормой. Согласно международной классификации выделяют следующие варианты медленноволновой активности:
- замедление основной активности;
- периодическое замедление;
- продолженное замедление.
Замедление основной активности констатируется, когда основные ритмы имеют более медленные частотные характеристики по сравнению с возрастной нормой: в 1 год — частота менее 5Гц, в 4 года — менее 6 Гц, в 5 лет — менее 7Гц, в 8 лет и старше — менее 8 Гц.
Периодическое замедление. Периодическое замедление может быть нерегулярным и ритмическим, генерализованным и региональным. Выраженное периодическое ритмическое генерализованное замедление (обычно с преобладанием в лобных отведениях) иногда наблюдается при генерализованных формах эпилепсии. Нерегулярное региональное замедление (чаще в височных отведениях) может быть косвенным ЭЭГ признаком парциальной эпилепсии или локального органического поражения головного мозга.
Продолженное замедление констатируют в случае если данный паттерн занимает около 90% записи, и нет реакции на внешние стимулы. Всегда является патологическим признаком и свидетельствует об прогрессирующем очагово-деструктивном поражении мозга. В этом случае медленно-волновая активность отражает изменение коркового электрогенеза, вызванного анатомическим дефектом неирональных сетей. Она может сочетаться с нормальным или замедленным основным ритмом; возникает по одному из отведений (например , левому височному) или по всей гемисфере. Как правило, представлено тета (реже дельта) активностью низкой амплитуды, не реагирующей на экзогенные стимулы.
4.4. Провоцирующие пробы
Провоцирующие пробы. Фоновая запись биоэлектрической активности головного мозга осуществляется в состоянии пассивного бодрствования пациента с закрытыми глазами. С целью выявления нарушений ЭЭГ применяются провоцирующие пробы, Наиболее значимые из них следующие:
- Открывание-закрывание глаз.
- Гипервентиляция.
- Ритмическая фотостимуляция.
- Фоностимуляция.
- Депривация сна.
- Стимуляция умственной активности.
- Стимуляция мануальной активности.
Подробно остановимся на первых.
Проба на открывание-закрывание глаз служит для установления контакта с пациентом. При этом медицинский работник убеждается, что пациент находится в сознании и выполняет инструкции. Данная проба позволяет выявить реактивность альфа-ритма и других видов активности на открывание глаз. В норме при открывании глаза блокируется альфа ритм, нормальная и условно нормальная медленноволновая активность. Наоборот, отсугсгвие реакции паттернов на открывание глаз — как правило, признак патологической активности. Блокирование при открывании глаз затылочной пик-волновой активности у пациентов с доброкачественной затылочной эпилепсией Гасто является важным дифференциальным признаком с симптоматической затылочной эпилепсией. Следует помнить, что при некоторых формах фотосенситивной эпилепсии, эпилепти-формная активность на ЭЭГ возникает в момент закрывания глаз. Это может быть связано с исчезновением фиксации взора при закрытых глазах. Данный феномен был описан Panayiotopoulos (1998 ) и назван им «fixation off» или фотосенситивностью.
Гипервентиляция реально проводима у детей после 3-х лет. Продолжительность от 3 мин у детей до 5 мин у взрослых. Гипервентиляцию нельзя проводить в самом конце записи ЭЭГ, так как патологическая активность нередко появляется спустя некоторое время после окончания пробы. Основное назначение гипервентиляции — выявление генерализованной пик-волновой активности, а иногда и визуализация самого приступа (обычно абсанса). Реже появляется региональная эпилептиформная активность. Согласно наблюдениям Благо-склоновой Н.К. и Новиковой Л. А. (1994 ), появление пароксизмальных вспышек медленных волн при гипервентиляции свойственно здоровым детям и подросткам и является вариантом нормы. По мнению Daly & Pediey (1997 ), патологическая реакция на гипервентиляцию включает только появление на ЭЭГ пик-волновой активности или выраженной асимметрии паттернов. Принципиально важно, что любая другая реакция, в том числе и появление дельта активности, — индивидуальный вариант нормы. Таким образом, согласно современным воззрениям, оценка пароксизмальной генерализованной (нередко с бифронтальным преобладанием) ритмической тета-дельта активности при гипервентиляции как гипотетической «дисфункции мезо-диэнцефальных структур» несостоятельна. Подобная оценка нормальных, по своей сути, паттернов не имеет никакого значения для клиники, приводит к терминологической путанице и напрасному беспокойству, как врачей неврологов, так и самих пациентов.
Ритмическая фотостимуляция (РФС ) является важнейшей пробой для выявления патологической активности при фотосенситивных формах эпилепсии. Используется классическая методика Jeavons & Harding (1975 ). Лампа стробоскопа должна находиться на расстоянии 30 см от закрытых глаз пациента. Необходимо использование широкого спектра частот, начиная от 1 вспышки в сек и, заканчивая частотой 50 Гц. Наиболее эффективна в выявлении эпилептиформной активности стандартная РФС с частотой 16 Гц. Возможны следующие реакции на РФС:
- Отсутствие очевидной реакции.
- Усвоение ритма: появление колебаний на ЭЭГ синхронно со вспышками при РФС.
- Фотомиоклонический ответ: при РФС возникает «трепетание » век и подергивание псриокулярной мускулатуры (миоклонический гиперкинез) синхронно со вспышками света. Это отражается на ЭЭГ отчетливым «ритмическим миографическим артефактом» в передних отведениях.
- Фотопароксизмалъный ответ: появление при РФС эпилептиформной активности, чаще возникают короткие разряды генерализованной быстрой (4 Гц и выше) полипик-волновой активности. По классификации фотосенситивных эпилепсии (Binnie и соавторы, 1992), выделяют форму фотосенситивной эпилепсии, при которой приступы возникают исключительно в лабораторных условиях во время РФС при ЭЭГ исследовании. Наиболее типично возникновение миоклонических пароксизмов, вовлекающих мышцы лица, плечевого пояса и рук, синхронно со вспышками света. В повседневной жизни приступы отсутствуют, даже при воздействии бытовых факторов РФС (мелькание света). Как правило, данная форма эпилепсии выявляется случайно при направлении больных на ЭЭГ исследование по причинам, не имеющим отношения к эпилепсии. Большинство авторов не рекомендуют назначение АЭП при этой форме, да и само отнесение таких случаев к эпилепсии вызывает сомнение. Фотопароксизмалъный ответ с высокой достоверностью коррелирует с наличием фотосенситивной эпилепсии.
4.5. Артефакты
Артефакты представляют собой любые графоэлементы на ЭЭГ, не являющиеся отражением электрической активности головного мозга. Подразделяются на механические и биоэлектрические. Механические артефакты бывают инструментальные, электродные и от электросети. Наиболее распространен артефакт вследствие «наводки » от сети переменного тока (отсутствие заземления, использование рядом различной медицинской аппаратуры) в виде появления синусоидальных колебаний частоты 50 Гц.
Биоэлектрические артефакты подразделяются на следующие:
- Миогенные артефакты. Наиболее частый вид артефактов. Регистрируются высокочастотные колебания миограммы, обычно преобладающие в височных отведениях. Они чаще вызваны напряжением жевательной мускулатуры, мимических мышц, мышц шеи. Артефакт от фасцикуляторного тремора напоминает аркообразный мю-ритм и наблюдается максимально в лобно-височных отведениях. Глоссокинетический артефакт возникает при ритмичных движениях языка, например, во время разговора или сосания (кормление младенцев во время ЭЭГ исследования).
- Сердечные и дыхательные артефакты. Электрод, находящийся на крупном сосуде, может вызывать артефакт, напоминающий графоэлемент реоэнцефалограммы. На ЭЭГ могут быть зарегистрированы электрокардиографические потенциалы, которые необходимо отличать от доброкачественных эпилептиформных нарушений детства. Данные потенциалы преобладают на электроэнцефалограмме при электроцеребральной инактивации (гибель мозга). Также встречаются артефакты, обусловленные экскурсией грудной клетки (часто во время проведения гипервентиляции).
- Окулографичесше артефакты. Связаны с активностью m. orbicularis oculi и регистрируются обычно в лобных отведениях. Возникают при ритмическом моргании (тикоидный гиперкинез), нистагме.
- Артефакты, вызванные изменением сопротивления кожи. Изменение сопротивления кожных покровов может быть обусловлено различными биохимическими процессами в организме. Наиболее часто данный тип артефактов возникает при волнении пациентов во время ЭЭГ исследования, что сопровождается выраженной потливостью.
При этом врач осуществляющий мониторинг должен уметь дифференцировать эти артефакты. Если, например, артефакт не уходит с течением времени, необходимо проверить электрод на полноценность соединения с усилителем, с пациентом и в случае необходимости заменить его/настроить.
5. Эпилептиформная активность
Эпилептиформная активность характеризуется появлением на ЭЭГ острых волн или пиков, которые резко отличаются от основной активности фона и возникают преимущественно у лиц, страдающих эпилепсией. Классификация нарушений ЭЭГ, принятая Американской ассоциацией нейрофизиологов [Daly & Pedley, 1997], придерживается строгой терминологии в обозначении патологических феноменов. В классификации общепринят термин «эпилептиформная активность», в связи с его исключительным применением к электроэнцефалографическим феноменам.
Согласно классификации нарушений ЭЭГ различают 9 межприступных (интериктальных ) и два приступных (иктальных ) эпилептиформных паттерна.
Межприступные эпилептиформные изменения:
- пики (спайки );
- острые волны;
- доброкачественные эпилептиформные нарушения детства (ДЭНД , «роландические » комплексы);
- комплексы пик-медленная волна;
- комплексы пик-медленная волна 3 Гц;
- медленные комплексы пик-медленная волна;
- множественные пики (полиспайки );
- гипсаритмия;
- фотопароксизмальный ответ (фотопароксизмальная реакция).
Иктальные эпилептиформные изменения:
- ЭЭГ приступа;
- ЭЭГ статуса.
Рассмотрим все указанные варианты эпилептиформных нарушений на ЭЭГ:
1. Пики (спайки ) — эпилептиформный феномен, отличный от основной активности и имеющий пикообразную форму. Период пика составляет от 40 до 80 мсек. Это специфичный эпилептиформный паттерн, который наблюдается в рамках различных форм эпилепсии (генерализованных и парциальных). Одиночные пики встречаются исключительно редко; обычно они предшествуют появлению волн. Согласно базисным принципам электрофизиологии, появление спайков на ЭЭГ отражает процессы возбуждения корковых нейронов, а медленных волн — процессы торможения.
2. Острая волна — эпилептиформный феномен, отличный от основной активности, период которого составляет 80-200 мсек. Согласно данным ряда авторов [Eed-Oloffson, 1971; Gregori 1993], этот паттерн редко наблюдается у здоровых людей, и высоко специфичен в отношении эпилепсии. Острые волны, также как и пики, могут регистрироваться в виде региональных, мультирегиональных и генерализованных феноменов. Острая волна может встречаться, как изолированно (особенно , при парциальных формах эпилепсии), так и предшествуя медленной волне. Следует помнить, что острые волны, как и пики, могут представлять нормальные физиологические феномены: доброкачественные эпилептиформные транзиты сна (BETS ), Wicket — потенциалы, 14 и 6 Гц позитивные спайки и некоторые другие.
3. Доброкачественные эпилептиформные нарушения детства (ДЭНД ) — эпилептиформный феномен, который представлен в виде стереотипного электрического диполя, состоящего из острой волны, с последующей медленной волной. Амплитуда негативного полюса составляет 150-300 мкВ, — часто в 2 раза больше позитивного. Общий период комплекса составляет 80-120 мсек. Этот паттерн легко узнаваем, благодаря типичной морфологии, напоминающей зубцы QRST на ЭКГ [Мухин К. Ю. и соавт., 2001]. Для комплексов ДЭНД характерна тенденция к их группированию (дуплеты , триплеты и т.д.), а также усиление их представленности и амплитуды в фазу медленного сна. Доброкачественные эпилептиформные нарушения детства возникают, преимущественно, в возрасте от 3 до 14 лет и являются характерным паттерном при идиопатических парциальных формах эпилепсии. С наступлением пубертата их выраженность уменьшается, и они в большинстве случаев постепенно исчезают после 14-15 лет. Предполагается, что данный ЭЭГ паттерн является возраст-зависимым и генетически детерминированным с аутосомно-доминантным наследованием с низкой пенетрантностью и вариабельной экспрессивностью [Stephani, 2002].
4. Комплексы пик-медленная волна — представляют паттерн, состоящий из пика и следующей за ним медленной волны. Наиболее часто комплексы пик-медленная волна регистрируются в виде генерализованных разрядов, представленность и амплитуда которых усиливается в ФМС, при ГВ и РФС. Данная ЭЭГ картина высоко специфична для идиопатических генерализованных форм эпилепсии детского и юношеского возраста. Однако по данным Doose & Baier (1987 ), в 10-17% случаев генерализованные комплексы пик-медленная волна могут быть обнаружены у клинически здоровых лиц, в основном, у родственников пробандов с абсансными формами эпилепсии.
В виде единичных паттернов комплексы пик-медленная (или острая-медленная) волна встречаются при криптогенных и симптоматических формах парциальной эпилепсии.
5. Комплексы пик-медленная волна с частотой 3 Гц — представляют регулярный разряд генерализованных паттернов, состоящих из единичных спайков со следующей медленной волной с частотой от 2,5 до 3,5 Гц. Согласно классификации нарушений ЭЭГ, для отнесения паттернов в данную группу, продолжительность этих комплексов должна составлять более 3 секунд. Частота комплексов во время разряда непостоянна. В начале разряда она составляет 3-4 Гц, тогда как к финалу снижается до 2,5-2,25 Гц. Характерно амплитудное преобладание паттернов в лобных отведениях. Медленный сон вызывает активацию пик-волновых комплексов. При этом продолжительность разрядов во время сна укорачивается и одновременно возможно некоторое замедление частоты комплексов. Данный ЭЭГ паттерн характерен для абсансных форм эпилепсии, особенно — детской абсанс эпилепсии. Продолжительность разряда пик-волновых комплексов более 3 сек с высокой вероятностью является иктальным феноменом типичных абсансов.
6. Медленные комплексы пик-медленная волна — представляют собой нерегулярные разряды комплексов пик (а чаще — острая волна)-медленная волна, частотой менее 2,5 Гц. Согласно классификации нарушений ЭЭГ, продолжительность этих комплексов должна составлять более 3 секунд. Комплексы состоят из би- и трифазных негативных острых волн с периодом 150-200 мсек. и следующих за ними высокоамплитудных (300 -400 мкВ) негативных медленных волн. Они билатерально-синхронны, однако, в ряде случаев возможна их амплитудная асимметрия и начальная асинхрония. Характерная особенность этого паттерна — тенденция к усилению выраженности изменений во время ФМС.
7. Полипики (множественные пики) — определяются как группа генерализованных билатерально-синхронных, следующих друг за другом 3-х и более пиков частоты от 10 Гц и выше. Каждая группа полипиков может завершаться медленной волной (полипик -волновые комплексы). Генерализованные полипики являются специфичным паттерном для миоклонических форм эпилепсии, таких как юношеская миоклоническая эпилепсия, доброкачественная миоклоническая эпилепсия младенчества. Однако этот паттерн может встречаться и при парциальных формах эпилепсии, у больных синдромом Леннокса-Гасто, а также в случаях прогрессирующих эпилепсии с миоклонусом (болезни Лафора, Унферрихта-Лундборга и др.).
8. Гипсаритмия — эпилептиформный паттерн, характеризующийся нерегулярной диффузной продолженной высокоамплитудной ( >300 мкВ) медленноволновой активностью (1 -3 Гц), на фоне которой регистрируются мультирегиональные пики и острые волны. В отдельных случаях возможно преходящее кратковременное уплощение этой активности (вплоть до биоэлектрического молчания). Данный вариант гипсаритмии Ohtahara (1978 ) назвал паттерном вспышка-угнетение. В некоторых случаях (симптоматический вариант синдрома Веста) гипсаритмия существенно доминирует в одной из гемисфер, сочетаясь со стойкими региональными спайками в этой зоне. Сон существенно модифицирует гипсаритмию: во время ФМС амплитуда и представленность эпилептиформных изменений увеличивается и становится периодической, тогда как в быстром сне она уменьшается или исчезает полностью.
9. Фотопароксизмальный ответ. Характеризуется появлением эпилептиформной активности, как генерализованного характера, так и регионального (преимущественно , в затылочных отделах коры головного мозга), которая появляется при предъявлении ритмической фотостимуляции различной частоты. Максимальный ответ наблюдается при частоте РФС около 16 Гц при закрытых глазах. Лучше выявляется при референтном монтаже. Фотопароксизмальный ответ может продолжаться и после окончания световой стимуляции, что характерно для фотосенситивных форм эпилепсии: первичная фотосенситивная эпилепсия, идиопатическая фотосенситивная затылочная эпилепсия, болезнь Унферрихта-Лундборга и некоторые другие.
10. Иктальные ЭЭГ паттерны. ЭЭГ приступа — внезапное изменение биоэлектрической активности, регионального или диффузного характера, ассоциированное с эпилептическим приступом. Во многих случаях для дифференциальной диагностики иктальных и межприступных эпилептиформных нарушений на ЭЭГ единственным методом является видео-ЭЭГ-мониторинг. Однако кратковременность возникновения некоторых паттернов (например , разрядов генерализованных пик-волновых комплексов, продолжительностью 1-2 сек) не всегда позволяет точно зафиксировать наличие приступа, синхронно с ним. В этих случаях ряд авторов рекомендует употреблять термин «субклинические эпилептиформные нарушения на ЭЭГ» [Binnie, 1993]. ЭЭГ паттерн приступа может возникать как генерализованно, так и регионально. Это высокоспецифичный феномен для эпилепсии, даже если он возникает без клинических симптомов. При наличии в клинике пароксизмов неясной этиологии этот паттерн доказывает эпилептическую их природу.
11. ЭЭГ статуса определяется в случае продолженных эпилептиформных паттернов ЭЭГ приступа или часто повторяющихся паттернов ЭЭГ приступа без восстановления нормального ритма фоновой записи между ними. Следует отметить, что ЭЭГ статуса может не коррелировать с клиническими симптомами эпилептического статуса. Классический тому пример — электрический эпилептический статус медленного сна; тяжелая форма эпилепсии с выраженными когнитивными нарушениями, при которой частота и выраженность эпилептических приступов может быть минимальна или приступы отсутствуют вовсе. Таким образом, даже высоко специфичные паттерны ЭЭГ приступа и ЭЭГ статуса, следует рассматривать только в контексте с клиническими данными. Особенности иктальной ЭЭГ при различных типах эпилептических приступов в рамках отдельных форм эпилепсии будут рассмотрены в следующих главах.
6. Расшифровка и заключение ЭЭГ
Таким образом мы подошли к интерпретации ЭЭГ-нарушений
Эти рекомендации не являются строгими правилами. Они относятся в первую очередь к стандартной ЭЭГ. При описании более специализированных записей (неонатальные записи, электроцеребральное молчание) представление технических деталей должно быть более полным — в соответствии со стандартами ACNS (1 — «Minimum Technical Requirements (MTR ) for Performing Clinical EEG»; 2 — «Minimal Technical Standards for Pediatric Electroencephalography»; 3 — «Minimum Technical Standards for EEG Recording in Suspected Cerebral Death»).
Отчет ВЭЭГ должен состоять из 3 основных частей:
- введение,
- описание,
- интерпретация, включая
- впечатление (мнение ) касательно нормальности или степени аномальности,
- корреляцию данных ЭЭГ с клинической картиной.
1. Введение.
Введение должно начинаться с описания специальной подготовки, если таковая предпринималась перед записью.
2. Описание.
Описание ЭЭГ должно включать все характеристики записи, включая нормальные и аномальные, представленные объективным способом, максимально избегая утверждений об их значимости.
Целью является полный и объективный отчет, который позволит другим ЭЭГ-специалистам придти к выводу касательно нормальности или степени аномальности записи по описанию — без необходимости просматривать исходную ЭЭГ. Этот вывод может отличаться от исходного вывода, поскольку в определенной мере субъективен.
Описание начинается с фоновой активности, доминирующей активности, ее частоты, количества (постоянная , преходящая), локализации, амплитуды, симметрии или асимметрии, является ли она ритмической или иррегулярной. Частота должна указывать в Гц или циклах в секунду. В целях стандартизации отчета рекомендуется определять амплитуду в отведениях, включающих смежные электроды по схеме 10-20. Желательно, но не обязательно, оценивать амплитуду в микровольтах. Это позволит избежать таких терминов, как «низкий », «средний » и «высокий ». Недоминирующая активность — частота, количество, амплитуда, локализация, симметрия или асимметрия, ритмичность или ее недостаточность, должна описываться с использованием таких же единиц измерения, как и для доминантной активности.
Если проводились тесты — должны быть описаны реакции на открытие и закрытие глаз, а также произвольных, целенаправленных движений. Включается описание указания на симметрию или асимметрию, полноту или неполноту, устойчивость или неустойчивость.
Аномальные записи, неонатальные записи, или записи только во время сна, могут не содержать четкого доминирующего ритма. В таких случаях, должны быть описаны другие виды активности (амплитуда , частота, и пр.) в любом порядке. Если запись демонстрирует заметную межполушарную асимметрию, характеристики по каждому полушарию должны быть представлены по отдельности (доминантная , недоминантная активность).
Вслед за фоновой активностью должно идти описание нарушений, не относящихся к фоновой активности. В описание включаются: тип нарушения (спайки , острые волны, медленные волны), распространенность (диффузные , локальные), топография или локализация, симметрия, синхронность (внутри — и межполушарная), амплитуда, временные характеристики (непрерывная , периодическая, эпизодическая, или пароксизмальная) и количество аномальных паттернов. Количество нарушений описывается субъективным способом, поскольку в клинической ЭЭГ невозможно точно измерить количество или соотношение.
Если аномалия является эпизодической, необходимо обратить внимание на отсутствие или наличие периодичности между эпизодами, ритмичность или иррегулярность паттерна внутри каждого эпизода. Необходимо представить временной диапазон продолжительности эпизодов.
В описании процедур активации необходимо включать утверждение об их качестве (например , хорошая, удовлетворительная или плохая гипервентиляция, длительность сна, стадии сна, которые были достигнуты во время исследования). Надо указать тип (глиссандо , пошаговая) фотостимуляции и диапазон частот стимуляции. Описываются эффекты гипервентиляции и фотостимуляции, включая нормальные и аномальные ответы. Если гипервентиляция или фотостимуляция не проводились, необходимо указать причину. Поскольку направляющий врач предполагает, что эти процедуры используются по умолчанию, он может ожидать описания их результатов — даже в том случае, если необходимость их проведения не была указана явно в направлении.
Нет нужды указывать на отсутствие определенных характеристик, за исключением нормальных, — таких как низкоамплитудная быстрая активность, сонные веретена, и др. Такие фразы как «отсутствие фокальной патологии» или «нет эпилептиформных нарушений» могут использоваться только в разделе интерпретации — при наличии явного или предполагаемого запроса направляющего доктора. Они не должны использоваться в описательной части.
Артефакты должны описываться только в сомнительных случаях (например , сохраняется вероятность того, что они отражают церебральную активность) или, когда они носят необычный характер, мешают интерпретации записи, а также когда они обладают определенной диагностической ценностью (например , миокимии, нистагм, и др.).
3. Интерпретация.
(I ) Впечатление — это субъективное мнение специалиста о степени нормальности записи. Описание записи предназначено, в первую очередь, для электроэнцефалографиста, который использует его для последующего вывода, или другого эксперта, и должно быть детальным и объективным. Впечатление, с другой стороны, пишется в первую очередь для направляющего врача, и, следовательно, должно быть по возможности сжатым. Большинство клиницистов из предшествующего опыта предполагают, что чтение детального описания не дает им существенно новой информации, и поэтому ограничиваются интерпретаций. Если оно слишком большое и выглядит иррелевантно клинической картине, клиницист может потерять интерес, что в итоге приводит в снижению пользы от всего ЭЭГ отчета. Если запись считается аномальной, желательно указывать ее степень — с целью облегчить сравнение между повторными исследованиями. Поскольку эта часть отчета носит довольно субъективный характер, степень нарушений может варьировать от лаборатории к лаборатории. Однако в каждой лаборатории следует четко определять критерии степени нарушения и строго следовать им.
После определения степени нарушений, необходимо указать причины, на основе которых строится вывод. Если присутствуют несколько видов нарушений, желательно ограничиться списком из двух или трех главных нарушений, которые наиболее характерны для данной записи. Если перечислять все нарушения, наиболее существенные «растворяются » в тексте и теряется значимость выводов. При наличии данных предыдущих ЭЭГ записей, необходимо включать их сравнение с результатами данного исследования.
(II ) Клиническая корреляция — это попытка показать насколько данные ЭЭГ укладываются (или нет) в общую клиническую картину. Оно может варьировать — в зависимости от того, кому оно адресуется. Для адресата, далекого от неврологии или ЭЭГ, оно должно быть более тщательным и выверенным.
Если ЭЭГ аномальная — это указывает на церебральную дисфункцию, поскольку ЭЭГ является отражением церебральной функции. Тем не менее, фраза «церебральная дисфункция» может звучать излишне угрожающе и должна использоваться только когда нарушение квалифицируется как «более чем легкое» и когда имеется достаточно клинической информации, чтобы считать такой вывод реалистичным в данном клиническом контексте. В остальных случаях допустимы предложения типа «Запись указывает на легкую иррегулярность церебральной функции». Определенные паттерны ЭЭГ являются подтверждающими для более или менее специфических клинических ситуаций; дельта фокус может говорить о структурном поражении в соответствующем клиническом контексте; определенные типы спайков или острых волн подтверждают потенциальный эпилептогенез. Если нарушение ЭЭГ соответствует клинической информации, которая содержит диагноз или подозрение на наличие подобного состояния, можно указать, что данные ЭЭГ согласуются или подтверждают диагноз.
Цифровые способы записи, генерации и передачи отчета позволяют, при необходимости, включать в отчет короткие отрезки реальной записи, в том числе с примерами нарушений.
7. ВЭЭГ-мониторинг в оценке эффективности противосудорожной терапии
Одним из основных критериев объективизации действия противоэпилептических препаратов является изменение биоэлектрической активности мозга, регистрируемое с помощью ЭЭГ.
Эти изменения носят различный характер и зависят от формы эпилепсии и применяемой терапии.
Помимо воздействия антиконвульсантов на эпилептическую активность, они также оказывают влияние на характер фоновой ритмической активности. Хорошо описаны изменения фоновой ритмики, появляющиеся при длительном приеме бензодиазепинов и барбитуратов.
При прогрессирующем течении заболевания отмечается нарастание индекса эпилептической активности в очаге.
Другим маркером отрицательной динамики является появление дополнительных очагов эпилептической активности. Они могут быть зависимы от первичного очага или существовать независимо.
К характеристикам проградиентного течения заболевания относится появление феномена вторичной билатеральной синхронизации (ВБС ).
К ЭЭГ-критериям, отражающим положительное влияние ПЭП, относятся: снижение индекса пароксизмальности в очаге, уменьшение количества эпилептических очагов и регресс эффекта ВБС.
Динамические ВЭЭГ-исследования в период отмены терапии с высокой точностью позволяют оценить риск возобновления приступов.
8. Эффективность ЭЭГ-мониторинга
Была проанализирована достоверность направляющего диагноза «Эпилепсия » у первичных пациентов, поступающих в эпилептологический стационар (НПЦ медицинской помощи детям, ДЗ Москвы).
Исследуемая группа составила 1154 пациента в возрасте от 0 до 18 лет. Всем пациентам проводились следующие методы обследования: оценка нервно-психического статуса, видео-ЭЭГ-мониторинг продолжительностью 6 и более часов, и, в большинстве случаев, МРТ головного мозга.
Результаты: Диагноз «эпилепсия » был достоверно подтвержден у 643 пациентов (56 %); у 240 (20 ,8%) пациентов не было получено электроэнцефалографических паттернов эпилепсии, но диагноз оставлен в прежнем виде с учетом данных анамнеза и других методов обследования; у 133 пациентов (11 ,5%) диагноз «эпилепсия » был снят; у 46 (4 %) пациентов диагностированы парасомнии; у 39 (3 ,4%) — псевдоэпилептические (психогенные ) приступы; у 8 (0 .7%) — тики; в группу пациентов из 45 (3 ,9%) человек вошли дети с аффект-респираторными пароксизмами, синдромом Туретта, синкопальными состояниями, хореоатетозом/дистонией, мигренью, аутизмом, синдромом Мюнхаузена, мастурбацией.
Таким образом, у 23,2% (267 ) пациентов диагноз «эпилепсия » был исключен. Наиболее частыми пароксизмальными состояниями, мимикрирующими эпилепсию являлись парасомнии и психогенные приступы. Нельзя также забывать о большой группе (11 ,5% — 133 пациента) абсолютно здоровых детей, ошибочная постановка диагноза «эпилепсия » у которых, в большинстве случаев, была связана с неверной трактовкой поведенческих реакций, характерных для того или иного возраста. В подавляющем большинстве этих случаев причины гипердиагностики эпилепсии заключались в недостаточно полном и точном сборе анамнеза, неправильной трактовке результатов ЭЭГ, а в ряде случаев в психологическом давлении родственников пациента на врача.
9. Заключение
Успешное лечение эпилепсии напрямую зависит от своевременно и правильно установленного диагноза. Использование неинформативных методов диагностики на стартовом этапе лечения эпилепсии приводит к трудностям в подборе адекватной терапии, прогрессированию заболевания. В ЭЭГ это проявляется в виде появления множественных вторичных очагов эпилептической активности, развитию феномена вторичной билатеральной синхронизации при фокальных формах и значительному нарастанию индекса генерализованных разрядов при генерализованных формах эпилепсии.
Нередко наличие у пациента эпилептических приступов, несмотря на их очевидную курабельность, побуждает врача необоснованно вводить социальные ограничения, применять полипрагмазию в лечении.
С другой стороны, необоснованная констатация ремиссии у пациентов с эпилепсией также имеет неблагоприятные для пациента последствия, поскольку сохраняются клинически «невидимые » виды приступов или эпилептиформная активность на ЭЭГ.
Отсутствие изменений в записанном фрагменте ЭЭГ бодрствования длительностью до 30 мин (рекомендации ILAE) может создать ложное впечатление о положительной динамике на фоне лечения. Опираясь на полученные данные, врач может ошибочно констатировать клинико-энцефалографическую ремиссию. С другой стороны, выявление эпилептической активности на контрольной динамической ЭЭГ на фоне подобранной терапии может содержать фрагмент эпилептической активности, которую врач ошибочно трактует как «отрицательную динамику». В некоторых случаях на коротких фрагментах записи ЭЭГ-характеристики могут выглядеть как «нормальные » при сохраняющихся приступах. При этом объективный анализ продолженной записи свидетельствует, что характер биоэлектрической активности у пациента значимо не менялся. Ошибки при интерпретации связаны с чередованием нормальных и патологических фрагментов ЭЭГ.
Можно утверждать, что объективная трактовка изменений ЭЭГ может проводиться только при проведении ВЭЭГ-мониторинга.
Введение в алгоритм диагностики и динамического обследования ВЭЭГ-мониторинга позволяет, используя объективные клинико-нейрофизиологические критерии, своевременно диагностировать заболевание, оценивать состояние больного на разных этапах лечения, оптимизировать терапевтическую тактику и избегать диагностических ошибок у пациентов с эпилепсиями и эпилептическими синдромами.
Анализ длительного катамнеза больных эпилепсией (взрослых и детей) позволил разработать и внедрить в специализированных отделениях и кабинетах высокодостоверный комплексный клинико-нейрофизиологический подход к дифференциальной диагностике эпилепсий и судорожных синдромов, значительно повысить качество проводимой терапии в этой сложной группе пациентов.
Врач-педиатр, врач высшей категории,
невролог Тамбиев И. Е..
Ковалев И. Г.